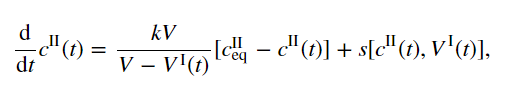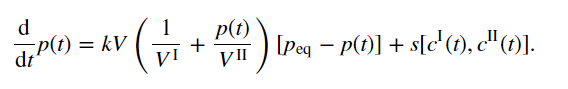活细胞中的冷凝物如何充当生物分子控制器?类似于调节房间温度的恒温器?这些相似之处依赖于凝聚物在动态变化条件下介导负反馈和稳定化学反应性质的能力。相分离系统中的反馈源于与不同浓度的外相共存的冷凝物。这种情况会引起通过冷凝物界面的分子通量,从而在内部和外部保持特定的浓度水平或保持浓度的函数。
研究领域:生物物理,细胞生物学,相分离
Christoph Weber、Christoph Zechner | 作者
赵雨亭 | 译者
邓一雪 | 编辑
在当下,细胞生物学的一个主要挑战仍然是解开细胞如何控制其生化反应周期的秘密。例如细胞是如何调节基因表达以应对压力?当资源稀缺时,它们的新陈代谢如何变化?控制理论已被证明有助于理解化学反应网络如何稳健地处理这些任务和其他任务[1]。这种方法中的基本要素是化学反馈回路,它创建类似于调节回路的控制机制,例如,加热温度系统、调节湿度或反应环境的pH值。
控制生化反应的理论主要集中在均匀、充分搅拌的环境上。然而,细胞内的大分子通常通过称为细胞器的特殊亚基在空间中高度组织。一些细胞器——例如细胞核——由膜结合。相比之下,另一类细胞器——生物分子凝聚物——显示出类液体液滴的标志性物理特性,它们为生化反应提供了化学上不同的环境[2–4]。
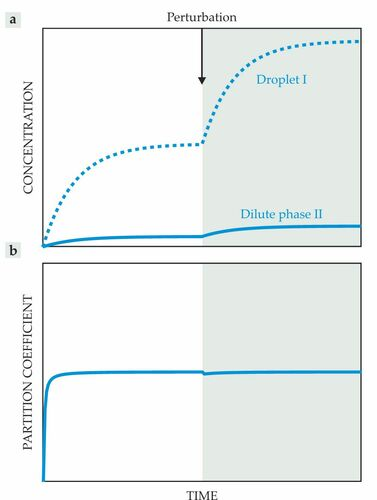
这种液滴可以作为活细胞中生化反应的微反应器(见下图)。它们的液体性质维持反应物的快速扩散,而它们的特定成分引起反应物在液滴内或外的分配。通常,冷凝物中反应物的浓度与外部的浓度不同。这些差异会改变反应通量,进而显着影响化学反应的反应产率和其他性质。这种经过修饰的通量如何控制细胞内的生物化学仍然知之甚少。
图1. 可以想象,液滴可以作为反馈控制器来调节细胞中的生化过程
在本文中,研究人员使用相分离物理学和控制理论来推测液体冷凝物如何在活细胞中实现反馈控制策略。
生化反应是生物学处理和控制细胞信息的基本构件。它们通过信号转导调节细胞代谢和环境线索的感知。同时,它们还调节基因的表达,其信息存储在DNA中,被转录并翻译成功能性蛋白质。
化学反应网络为这些过程提供了有用的数学描述。在这种形式中,分子被描述为粒子,它们在反应体积内随机扩散。当两个(或多个)正确类型的分子彼此相遇时,它们会发生化学反应。一旦反应发生,原来的反应物分子就转化为一组产物分子。在双分子反应中,A+B⇋C+D,分子类型A和B转化为C和D。细胞内过程通常包括多个反应循环,其中一个反应的产物作为另一个反应的反应物,等等。这种相互依赖导致了复杂的化学反应网络,这些网络可以表现出丰富而复杂的动力学行为,例如振荡或多重稳定性。
化学反应网络的动力学可以使用化学动力学理论来描述,该理论捕捉不同化学相互作用分子的浓度如何随时间变化。在其传统形式中,化学动力学适用于混合良好的系统,即运行速度比分子在反应体积内扩散慢得多的反应。换句话说,反应体积的大小远小于相应的反应扩散长度尺度。因此,由反应引起的局部浓度变化将通过快速扩散立即均匀化。在这样一个混合良好的系统中,反应发生的速率仅取决于系统中存在的分子数量;这些分子的确切空间位置变得无关紧要。可以根据反应物浓度水平的时间动态来充分表征混合良好的化学系统。
细胞内生化网络的一个标志是它们对分子成分的随机波动、周围环境的变化或其他潜在干扰的显著鲁棒性。先前的研究表明,负反馈机制在稳定各种细胞过程以抵抗此类干扰方面发挥着关键作用。蛋白质抑制自身表达的自调节基因网络是典型的例子[5]。如果蛋白质水平低于某个设定点,蛋白质对转录的抑制作用减弱,蛋白质水平开始上升。随着接近(或超过)设定点,负反馈变得更加明显,进而减少更多蛋白质的产生。通过这种方式,蛋白质水平可以保持在狭窄的范围内,尽管可能会影响系统的潜在干扰。
尽管这个简单的例子说明了负反馈调节背后的核心思想,但生物系统通常要复杂得多,涉及许多反应循环、并发反馈循环和强非线性。提取阐明此类系统功能的简单原理变得具有挑战性。
控制理论为抽象和分析受反馈影响的动态系统提供了丰富的框架。除了在工程中的传统应用之外,控制理论已被证明是研究和逆向工程生物系统的有效方法。事实上,通过在控制理论的形式主义中构建给定的生物系统,大量的数学工具和概念变得适用。这些工具和概念使得评估所考虑系统的基本属性成为可能,即使其某些细节未知。
在经典控制问题中,研究人员考虑一个任意的物理过程,其中需要输入x和输出y。例如,y可以对应于房间的温度,而x可以是该房间中加热设备的功率。在此基础上,目标是通过负反馈回路相对于某个参考值u(例如所需的环境温度)来控制输出y。该循环包含一个控制器,它测量y和u之间的失配并计算适当的控制输入x以减少失配。
将y和u映射到x的函数称为控制律。在工程应用中,它是根据性能和可用资源来选择的。最简单的控制法则之一称为比例控制。在这里,控制器纯粹根据y和u之间的瞬时失配来调整x。更准确地说,x=G(u−y)=eG,其中G是反馈增益,它决定了控制器对失配e的反应强度。
当G足够高时,闭环动力学将主要由G(u−y)控制,这反过来将有效地减少u和y之间的失配。即使存在潜在干扰d也能进行控制;这种扰动可能是对系统的物理扰动,也可能是额外的、可能未知的通量和波动。
通过扩展控制律并添加与失配e的积分、其导数或两者成正比的项,可以实现更有效的控制器。与纯比例控制器相比,所谓的比例积分微分控制器表现出改进的动态特性,例如零稳态误差或向所需参考值u的改进收敛。
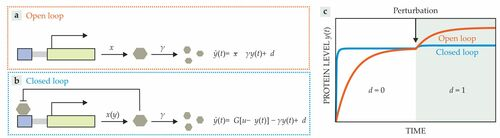
为了更具体地展示如何使用控制理论研究生化反馈系统,让研究人员考虑一个简单的蛋白质表达玩具模型。在该系统中,蛋白质以速率常数x和γ产生和降解(见图2a)[6]。
动力学受到扰动d的影响,为简单起见,研究人员将其视为常数。在稳定状态下,蛋白质水平由y=(x+d)/γ给出。也就是说,蛋白质水平y对扰动d很敏感,并与它成线性关系。
现在考虑对网络的修改:该蛋白质通过与其自身的启动子结合来负调节其他蛋白质的表达(图2b)。在这种情况下,可以使用Hill型函数描述蛋白质生产率,使得x(y)=λKn/(Kn+yn);n、K和λ是正参数。对于小y,生产率将接近λ,而对于大y,它接近于零。围绕y的稳态值对函数进行线性化揭示了一个简单的控制器结构,其中蛋白质生产速率大约由某些参考u和蛋白质水平y之间的不匹配给出,乘以恒定增益G(见图2b)。u和G都是动力学参数K、n和λ的函数;为了紧凑,这里省略了显式依赖。
这个简单的分析表明,研究人员的遗传反馈电路的作用,就像一个比例反馈控制器一样,它试图将蛋白质水平y维持在某个目标值u。因此,如果G足够大,即使存在扰动d,y也会达到接近u的值。当比较受开环和闭环遗传电路不断扰动的时间的蛋白质水平时,可以看出这一点(图2c)。虽然开环电路对扰动很敏感,但它们在相应的闭环系统中受到很大程度的抑制。参考文献[6-8]提供了关于控制理论如何阐明生物和其他物理系统的更多信息。
液体冷凝物可以通过相分离形成多组分混合物,并在较低浓度的环境中稳定共存。这种混合物的热力学行为受自由能的最小化控制,这解释了相互作用能和熵之间的竞争[9]。为了发生相分离,溶剂中的分子需要相互吸引或排斥溶剂分子,这样共存状态的相互作用能的增益超过了形成界面的相应缺点。
这种相共存受到各种物理化学控制参数的影响,例如分子和盐的浓度、pH值和温度。参数定义了相图,它描述了共存相的平衡参数,即冷凝物及其环境。在分子的固定平均浓度下自由能的最小化意味着共存相在热力学平衡时具有相同的化学势。化学势对应于自由能密度的斜率。最重要的是,共存相可能不仅在各自的浓度上不同,而且在盐浓度、pH值和其他控制参数上也可能不同。
空间异质化学势导致分子通量,其存在表明系统未处于热力学平衡。由许多液滴组成的乳液就是一个例子。要了解为什么乳液不处于平衡状态,研究人员需要考虑液滴界面。在单个液滴的情况下,总界面面积最低,因此自由能最小。因此,许多液滴的乳液随时间演变为对应于热力学平衡的单个液滴。
化学反应会影响相的共存。在热力学平衡时,化学反应对系统施加了进一步的约束:反应方案每一侧的反应物化学势总和(由化学计量因素加权)必须平衡。
一般而言,在热力学平衡状态下,在具有M个独立成分和s个化学反应的混合物中,只有(M−s)个独立浓度[10]。其余浓度通常与反应的守恒量有关,例如总质量或反应物的总分子体积。例如,具有可逆化学反应A⇋B(s=1)的不可压缩三元混合物(M=2)给出一个独立的浓度,该浓度等于化学反应的守恒量。通常,相图与没有化学反应的相同系统相比显示出较低的维度,并且反应的相应相图的轴——共存相的最大数量——对于每个化学约束减少一个。因此,根据吉布斯相规则,热力学平衡时的化学反应会抑制多相的共存。减少到一组较小的守恒变量和抑制多相共存不一定会远离热力学平衡。
一个有趣的例子是通过燃料分子的供应维持远离平衡的化学反应。这些分子在反应步骤中不可逆转地降解,破坏了反应速率的详细平衡,并导致在热力学平衡下不可能出现的现象。这种反应的一个范例是液体冷凝物的分裂,它代表了热力学系统的逆动力学变为通过融合更加异质[9]。
共存相也会影响化学反应。要了解原因,请考虑两种分子的简单混合物:支架和内容物。支架分子构成冷凝物,而内容物进行化学反应。如果内容物是稀释的,则它们对富含支架的冷凝物的影响可以忽略不计[11]。
然而,由于冷凝物的存在,做出反应的客户端会经历空间异构环境。这种环境本质上对化学反应有两个关键影响。首先,反应客户被分配进或出冷凝物,其中分配来自客户和支架分子之间的相互作用。其次,在每个阶段内,客户遵循反应扩散类型的动力学,其反应速率特定于每个阶段。速率出现是因为每个相的浓度和反应速率常数不同。这两种效应有助于液体冷凝物对化学反应的调节,它们体现了冷凝物作为生化反应控制策略的潜力。
为了更好地理解相分离系统如何控制反应,从控制理论的角度分析它们的动力学特性是有用的。为此,研究人员考虑了一个简单的相分离系统,该系统分为两相:一相富含支架分子,另一相缺乏支架分子(图3a)。
支架浓度的动力学方程可以从热力学考虑中推导出来,与一级反应速率常数相比,通过液滴的扩散速度较快。在该限制下,每个相中的浓度近似均匀,这意味着稀相支架浓度cII(t)可以用常微分方程描述[12]:
例如,其中k是取决于扩散率的平衡弛豫率。术语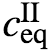 是平衡稀相浓度,V和VI(t)分别是总体积和液滴体积。该方程捕获了稀相和密相之间支架分子的分配。cII(t)的动力学受反应通量s的影响,可能是通过支架分子的产生和周转。该反应通量驱使系统远离平衡。
研究人员现在可以使用反馈控制的概念重铸这个相分离系统。特别是,如果研究人员将稀相支架浓度视为系统的输出y(t),那么右侧的两个交换通量的总和实现了可以认为是当前浓度之间的误差计算——即输出y(t)=cII(t)——以及相应的平衡浓度,或参考值
是平衡稀相浓度,V和VI(t)分别是总体积和液滴体积。该方程捕获了稀相和密相之间支架分子的分配。cII(t)的动力学受反应通量s的影响,可能是通过支架分子的产生和周转。该反应通量驱使系统远离平衡。
研究人员现在可以使用反馈控制的概念重铸这个相分离系统。特别是,如果研究人员将稀相支架浓度视为系统的输出y(t),那么右侧的两个交换通量的总和实现了可以认为是当前浓度之间的误差计算——即输出y(t)=cII(t)——以及相应的平衡浓度,或参考值 。所得误差乘以时间相关增益G=kV/[V–VI(t)]。因此,尽管存在额外的通量和扰动,液滴仍像比例控制器一样将稀相支架浓度保持在某个设定点(见图3b)。
控制器的有效性取决于反馈增益G,它来自混合物的物理特性,例如支架分子之间的相互作用强度及其扩散率。
对于分配为液体冷凝物的稀释分子,也可以确定类似的反馈控制结构。然而,与支架分子相比,控制器不再调节绝对浓度,而是调节内部和外部浓度之间的比例。研究人员称该比率为有效分配系数p(t)=cI(t)/cII(t)。p(t)的变化可以用等式表示
peq(定义为
。所得误差乘以时间相关增益G=kV/[V–VI(t)]。因此,尽管存在额外的通量和扰动,液滴仍像比例控制器一样将稀相支架浓度保持在某个设定点(见图3b)。
控制器的有效性取决于反馈增益G,它来自混合物的物理特性,例如支架分子之间的相互作用强度及其扩散率。
对于分配为液体冷凝物的稀释分子,也可以确定类似的反馈控制结构。然而,与支架分子相比,控制器不再调节绝对浓度,而是调节内部和外部浓度之间的比例。研究人员称该比率为有效分配系数p(t)=cI(t)/cII(t)。p(t)的变化可以用等式表示
peq(定义为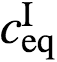 /)是平衡时的分配系数,VI和VII分别是稀相和密相的体积。该表达式表明,对于内容物分子,液滴类似于具有输出y(t)=p(t)、参考u=peq和反馈增益的比例控制器(见图4a)。因此,虽然每个阶段的客户集中度对额外的通量和扰动很敏感,但分配系数可以通过反馈控制回路稳健地保持(见图4b)。反过来,反馈提供了有趣的方法来控制下游化学过程(例如酶促反应)的浓度。
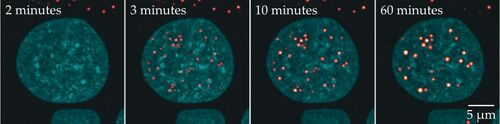
生物分子凝聚物作为细胞内反馈控制器的想法有多合理?尽管该领域还知之甚少,但最近的几项研究指向了生物系统,其中液滴介导的反馈控制可能确实相关。一个例子是抑制浓度波动[12-14]。
由于冷凝液内外支架分子的浓度在热力学上受到限制,因此预计冷凝液会通过改变其大小来响应波动的浓度。每个相内的浓度受到的影响要小得多。与图3b和3c中所示的支架控制方案一致,因此可以将冷凝物理解为反馈控制器,它试图最小化每个相中的浓度与其参考平衡值之间的不匹配。
控制器的有效性(由反馈增益G反映)取决于基础物理参数和相互作用。早期的研究使用控制理论概念来确定同质生化反馈回路中噪声抑制的硬下界。了解空间划分如何影响这些结果是一个重要的开放性问题[15]。
最近在转录凝聚物的背景下提出了一种相关且更复杂的负反馈回路。人们认为它们通过集中所需的转录机制(如转录因子、辅助因子及聚合酶)来增强信使RNA (mRNA) 的转录[16]。
有趣的是,该研究发现,一旦新转录的mRNA超过某个设定点,就会促进凝聚物的溶解,从而阻止转录。这产生了一个负反馈通路,尽管存在潜在的干扰,它可以可靠地控制转录的持续时间和输出。
越来越多的证据表明,冷凝物可以控制聚集过程,包括细胞骨架基础分子(如肌动蛋白和微管)的形成,以及与疾病相关的原纤维(如淀粉样蛋白)的形成。尽管聚集体的生物学作用多种多样,但它们的形成具有一些共同的物理原理。初始聚集体通过初级成核形成并主要在其末端生长,次级成核允许在现有聚集体表面附近形成小聚集体[17]。
有趣的是,凝聚物可以调节已经仅通过初级成核形成的聚集体的反馈机制。随着单体分配到冷凝液中并在其中聚集,它们在冷凝液中的浓度应该会降低。
然而,一部分聚集的单体被分配通量补充,它试图保持单体在冷凝物内部和外部的分配。因此,有利于凝聚物内部聚集体的生长,而外部聚集体受到抑制。对于这个简单的聚合过程,冷凝物提供了一种让人联想到控制电路的机制。富含支架的冷凝物作为控制器,调节易聚集单体的分配,从而调节成核动力学。该反馈类似于图4和等式2中描述的客户端示例。
研究人员最近发现了一种类似但稍微复杂一些的反馈机制,该机制由凝聚物介导。它发生在经历二次成核并在其末端生长的原纤维系统中。有趣的是,该机制通过改变冷凝物的特征物理参数(例如单体分配和冷凝物尺寸)来控制聚集体的数量和大小[18]。这些发现支持冷凝物在调节生理组装或与疾病相关的淀粉样蛋白原纤维方面的潜在相关性。
/)是平衡时的分配系数,VI和VII分别是稀相和密相的体积。该表达式表明,对于内容物分子,液滴类似于具有输出y(t)=p(t)、参考u=peq和反馈增益的比例控制器(见图4a)。因此,虽然每个阶段的客户集中度对额外的通量和扰动很敏感,但分配系数可以通过反馈控制回路稳健地保持(见图4b)。反过来,反馈提供了有趣的方法来控制下游化学过程(例如酶促反应)的浓度。
生物分子凝聚物作为细胞内反馈控制器的想法有多合理?尽管该领域还知之甚少,但最近的几项研究指向了生物系统,其中液滴介导的反馈控制可能确实相关。一个例子是抑制浓度波动[12-14]。
由于冷凝液内外支架分子的浓度在热力学上受到限制,因此预计冷凝液会通过改变其大小来响应波动的浓度。每个相内的浓度受到的影响要小得多。与图3b和3c中所示的支架控制方案一致,因此可以将冷凝物理解为反馈控制器,它试图最小化每个相中的浓度与其参考平衡值之间的不匹配。
控制器的有效性(由反馈增益G反映)取决于基础物理参数和相互作用。早期的研究使用控制理论概念来确定同质生化反馈回路中噪声抑制的硬下界。了解空间划分如何影响这些结果是一个重要的开放性问题[15]。
最近在转录凝聚物的背景下提出了一种相关且更复杂的负反馈回路。人们认为它们通过集中所需的转录机制(如转录因子、辅助因子及聚合酶)来增强信使RNA (mRNA) 的转录[16]。
有趣的是,该研究发现,一旦新转录的mRNA超过某个设定点,就会促进凝聚物的溶解,从而阻止转录。这产生了一个负反馈通路,尽管存在潜在的干扰,它可以可靠地控制转录的持续时间和输出。
越来越多的证据表明,冷凝物可以控制聚集过程,包括细胞骨架基础分子(如肌动蛋白和微管)的形成,以及与疾病相关的原纤维(如淀粉样蛋白)的形成。尽管聚集体的生物学作用多种多样,但它们的形成具有一些共同的物理原理。初始聚集体通过初级成核形成并主要在其末端生长,次级成核允许在现有聚集体表面附近形成小聚集体[17]。
有趣的是,凝聚物可以调节已经仅通过初级成核形成的聚集体的反馈机制。随着单体分配到冷凝液中并在其中聚集,它们在冷凝液中的浓度应该会降低。
然而,一部分聚集的单体被分配通量补充,它试图保持单体在冷凝物内部和外部的分配。因此,有利于凝聚物内部聚集体的生长,而外部聚集体受到抑制。对于这个简单的聚合过程,冷凝物提供了一种让人联想到控制电路的机制。富含支架的冷凝物作为控制器,调节易聚集单体的分配,从而调节成核动力学。该反馈类似于图4和等式2中描述的客户端示例。
研究人员最近发现了一种类似但稍微复杂一些的反馈机制,该机制由凝聚物介导。它发生在经历二次成核并在其末端生长的原纤维系统中。有趣的是,该机制通过改变冷凝物的特征物理参数(例如单体分配和冷凝物尺寸)来控制聚集体的数量和大小[18]。这些发现支持冷凝物在调节生理组装或与疾病相关的淀粉样蛋白原纤维方面的潜在相关性。
本文概述的想法说明了活细胞中的冷凝物如何充当生物分子控制器,类似于调节房间温度的恒温器。这些相似之处依赖于凝聚物在动态变化条件下介导负反馈和稳定化学反应性质的能力。相分离系统中的反馈源于与不同浓度的外相共存的冷凝物。这种情况会引起通过冷凝物界面的分子通量,从而在内部和外部保持特定的浓度水平或保持浓度的函数。
研究人员将研究人员的生物分子示例限制为比例控制,这是最简单的反馈控制架构之一。然而,当相分离与额外的反应循环相结合时,可以实现更稳健和有效的控制策略,例如积分反馈控制[1,8]。这种可能性为进一步研究提供了一个富有成效的途径。
但是,为什么认为冷凝物可以调节反馈控制的观点首先是有帮助的呢?首先,考虑一个不同的问题:通过对每个晶体管中的电子传输物理建模来描述现代计算机有用吗?对于许多物理学家来说,这个问题的答案是否定的——因为这样的方法会非常复杂,并且可能无法捕捉到系统的涌现特性。
细胞承载着极其复杂的生化过程网络,其中许多研究人员才刚刚开始了解。通过控制理论分析此类系统提供了一种通用策略来提取生物过程的相关结构和动态特征,而无需对系统进行详细的物理描述。通过控制理论了解空间区室化和生化反应之间的相互作用可能为理解甚至控制活细胞的功能开辟新的途径。
1.T.-M. Yi et al., Proc. Natl. Acad. Sci. USA 97, 4649 (2000).
2.A. A. Hyman, C. A. Weber, F. Jülicher, Annu. Rev. Cell Dev. Biol. 30, 39 (2014).
3.Y. Shin, C. P. Brangwynne, Science 357, eaaf4382 (2017).
4.S. F. Banani et al., Nat. Rev. Mol. Cell Biol. 18, 285 (2017).
5.Y. Dublanche et al., Mol. Syst. Biol. 2, 41 (2006).
6.D. Del Vecchio, A. J. Dy, Y. Qian, J. R. Soc. Interface 13, 20160380 (2016).
7.J. Bechhoefer, Rev. Mod. Phys. 77, 783 (2005).
8.M. Khammash, M. Di Bernardo, D. Di Bernardo, in 2019 IEEE 58th Conference on Decision and Control (CDC), IEEE (2019), p. 916. Google ScholarCrossref
9.C. A. Weber et al., Rep. Prog. Phys. 82, 064601 (2019).
10.O. Adame-Arana et al., Biophys. J. 119, 1590 (2020).
11.S. F. Banani et al., Cell 166, 651 (2016).
12.A. Klosin et al., Science 367, 464 (2020).
13.T. Stoeger, N. Battich, L. Pelkmans, Cell 164, 1151 (2016).
14.D. Deviri, S. A. Safran, www.biorxiv.org/content/10.1101/2021.01.05.425486v1.
15.I. Lestas, G. Vinnicombe, J. Paulsson, Nature 467, 174 (2010).
16.J. E. Henninger et al., Cell 184, 207 (2021).
17.T. C. T. Michaels et al., Annu. Rev. Phys. Chem. 69, 273 (2018).
18.C. Weber, T. Michaels, L. Mahadevan, eLife 8, e42315 (2019).
https://physicstoday.scitation.org/doi/10.1063/PT.3.4771
集智斑图顶刊论文速递栏目上线以来,持续收录来自Nature、Science等顶刊的最新论文,追踪复杂系统、网络科学、计算社会科学等领域的前沿进展。现在正式推出订阅功能,每周通过微信服务号「集智斑图」推送论文信息。扫描下方二维码即可一键订阅:

点击“阅读原文”,追踪复杂科学顶刊论文