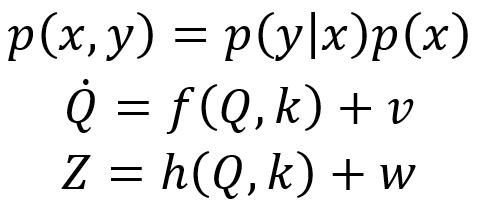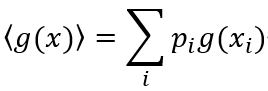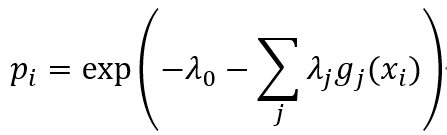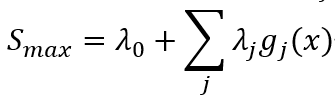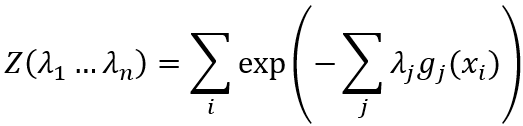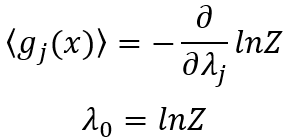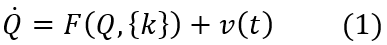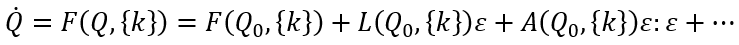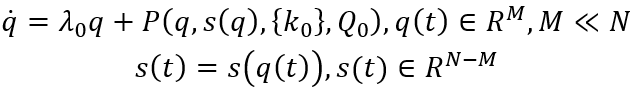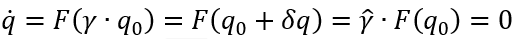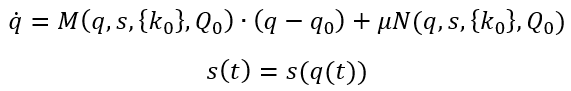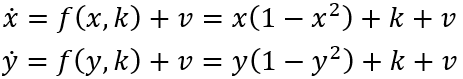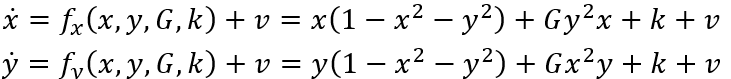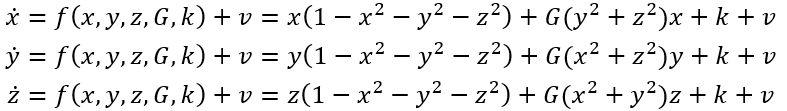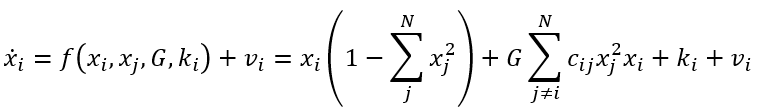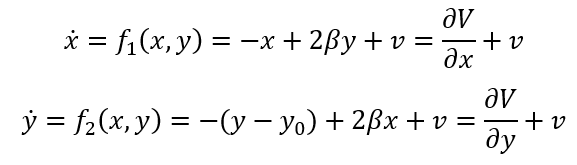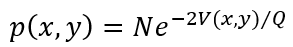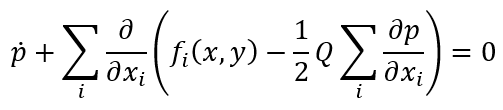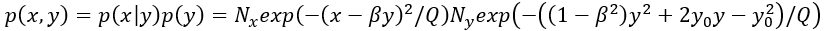我们的大脑在一定程度上是贝叶斯推理系统,生成内部模型对外部世界作出预测,然后将预测与感官输入不断地进行对比,形成预测误差并更新内部模型。2022年初发表于跨学科期刊 Journal of Physics: Complexity 的一项研究提出一种神经机制——大脑从脑网络连接的对称性破缺中生成内部模型。涌现的观点说明了自由能如何与内部模型联系起来,以及如何由神经底层中产生。研究关注在信息论框架内整合确定性过程和随机性过程,从而将信息熵和自由能与脑网络中的涌现动力学机制和自组织机制联系起来。
人类大脑是由数以百亿计的神经元相互连接所构成的复杂系统,吸引来自不同领域的无数科学家从不同尺度探究其中奥秘——从解析神经元之间的相互作用机理,到刻画皮层柱之间的连接形式,再到探究脑区间不同认知功能的分离与整合模式,以期揭示人脑这一最为复杂的神经系统的工作模式,理解语言、学习、社会交往等高级认知活动的底层神经机制,并有望启发通用人工智能机器人的设计。为探讨认知、智能、意识等根本性问题,集智俱乐部组织了「神经动力学模型」、「计算神经科学」、「NeuroAI」等系列主题读书会,并将在春节后开启「自由能原理与强化学习」读书会和更多活动探讨意识的量化与建模问题,探索神经科学、系统科学、生物学、物理学与人工智能如何相互启发,欢迎感兴趣的朋友持续关注。
研究领域:脑网络,对称性破缺,神经动力学,信息熵,热力学,贝叶斯大脑,自由能原理,自组织,涌现
Viktor Jirsa, Hiba Sheheitli | 作者
JawDrin | 译者
彭菘峻 | 审校
论文题目:
Entropy, Free Energy, Symmetry and Dynamics in the Brain
https://iopscience.iop.org/article/10.1088/2632-072X/ac4bec/meta
1. 引言
2. 大脑的信息论框架——系统演化建模
3. 自组织系统中的涌现——系统动力学建模
4. 大脑中的等变动力学
摘要:神经科学是根植于各种领域的概念及理论的家园,包括信息论、动力系统理论和认知心理学。但并非所有的领域都可以连贯地联系起来,有些概念是不可通约的,而且特定领域的术语给整合带来障碍。尽管如此,概念整合仍然是提供直觉和巩固的理解形式,没有概念整合,进展将是无头苍蝇。本文关注在信息论框架内整合确定性过程和随机性过程,从而将信息熵和自由能与脑网络中的涌现动力学机制和自组织机制联系起来。我们确定了神经元群(neuronal populations)的基本属性,该属性导致等变矩阵出现在网络中,而在该网络中,复杂行为可以通过流形上的结构化流来表示,从而建立与脑功能理论相关的内部模型。我们提出一种从脑网络连接的对称性破缺中生产内部模型的神经机制。涌现的观点说明了自由能如何与内部模型联系起来,以及它们如何由神经底层中产生。
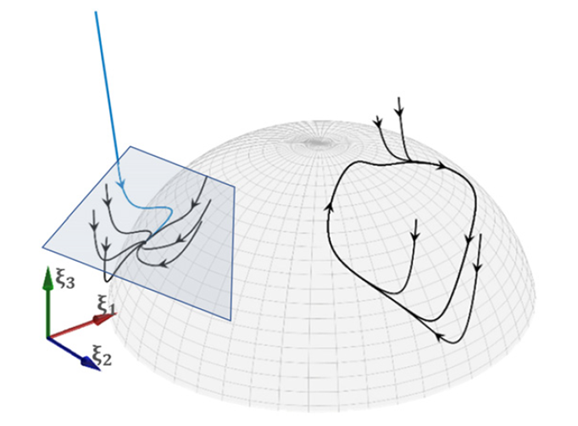
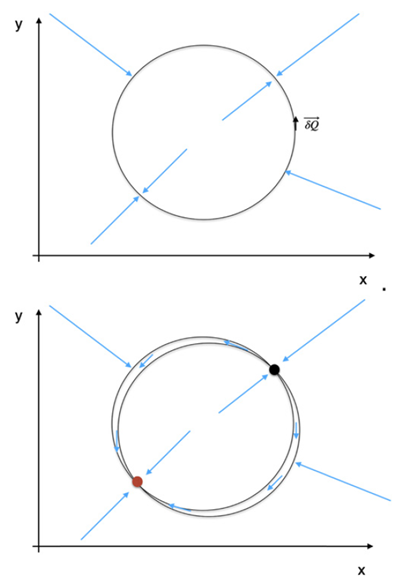
预测编码(Predictive coding)是当代最具影响力的脑功能理论之一[1-3]。该理论基于大脑作为贝叶斯推理系统运行这一直觉,如此实现内部生成模型对外部世界的预测。这些预测不断地与感官输入进行对比,形成预测误差并更新内部模型(见图1)。从理论的角度来看,用预测编码的形式表示脑功能是如此迷人,因为这解决了不同领域过多的深奥概念。这样便有机会将抽象概念联系进一个整合框架内,这些抽象概念如动力学、确定性作用和随机性作用、涌现、自组织[4-6]、信息、熵、自由能[7,8]、稳态等等。该跨领域的整合符合对这些复杂抽象概念的直观理解,尽管通常为保留易处理性,对于给定概念的所有复杂性层面没有全部被等价地刻画。例如,在预测编码理论中使用简单的内部模型(如在做决策时,动力学被简化为转换),但是简单的模型很难推广到更复杂的行为。当焦点在过程的推理部分时,这种做法理所当然,而我们不希望这样做。实际上我们强调内部模型在大脑激活方面的神经基础以及要与复杂行为涌现的理论相联系。不过,考虑到信息论概念(尤其是熵和自由能)的重要性,在预测编码方面,这样的尝试要求它们与当代脑网络模型中存在的概率分布函数和相关的确定性作用和随机性作用进行整合(相关观点同见[9-11])。
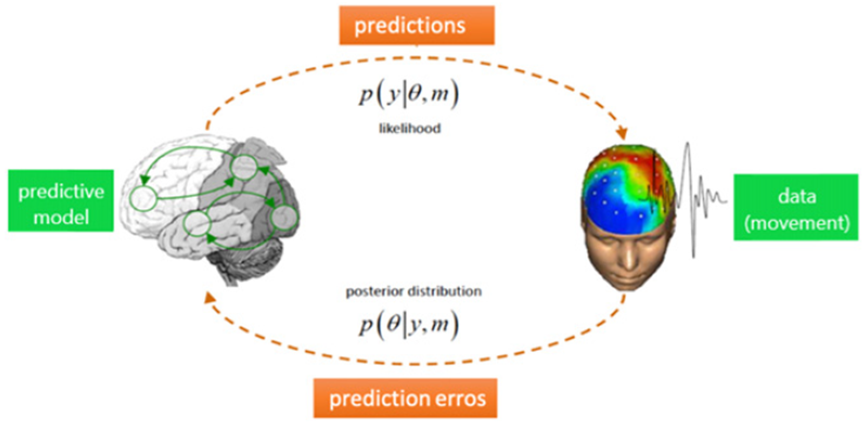
图1:贝叶斯大脑假说(Bayesian brain hypothesis)的推理过程图示。图左侧的生成模型表现为内部神经动力学,通过感知和行动(图右侧)与外部世界进行信息交换来实现模型更新。
Karl Friston首次提出把自由能作为大脑功能的一个原则[12-14],从数学上阐述了自适应、自组织系统如何抵抗自然的(热力学的)无序倾向。随着时间推移,自由能原理已经从赫姆霍兹机(Helmholtz machine)中使用的自由能概念里发展出来,在预测编码背景下用来解释大脑皮层反应,并逐渐发展为智能体的一般原则,这也被称为主动推理(Active Inference)[15]。贝叶斯推理过程和最大信息原理(maximum information principle)两者实际上都可重新阐述为自由能最小化问题。尽管这些关于自由能的概念在两个相关的框架中都有使用,但它们并不完全等价。模棱两可或因为这样的事实,即它们的一般形式类似于热力学亥姆霍兹自由能(Helmholtz free energy),但是却来自于两种不同的推理路线(详见[15])。第一个是所谓的“来自约束的自由能”,对应于最大信息原理下最小化的自由能,表示确定性约束和随机作用之间的权衡[7]。我们主要考虑的便是这种类型的自由能及其约束。另一个是变分自由能(variational free energy)并与贝叶斯大脑假说有关。这种自由能概念源于对贝叶斯规则的重新表述,即表述为寻找最小化相对熵(KL-散度)的概率分布这一优化问题,其中相对熵表示偏离精确贝叶斯后验的误差。
>> 关注2024年最新「自由能原理与强化学习」读书会:
确定约束在流形上的结构化流(structured flows on manifolds, SFMs)[16,17]框架中以动力学的形式表示,其中结构化流属于网络中产生的低维动力系统,因此是大脑理论中表示内部模型的主要候选者。通过概率分布的介导,经验可得的函数(如放电率、能量、方差等等)上的相关性都含有自由能和SFMs两者之间的联系,其中,概率分布在系统中确定性和随机性作用的相互作用下形成。Ilya Prigogine 在阐述熵的含义时,详细说明了这些作用之间的紧密联系。在这里,时间(的概念)超越了重复和退化的概念,达到建设性的不可逆性概念,如生命系统所表现的那样,通过与其所处的环境进行熵的交换来永存自己。生物学被认为需要将不可逆的时间刻在物质之上。在神经科学的背景下,这让我们想起Ingvar的假定,即大脑有能力通过其时间极化结构模拟“将来的记忆”[20],以及大脑能维持和导航分布在不同脑区的过去、现在和将来的经验。
尽管熵的数学公式最先出现在经典热力学(涉及诸如热、温度和能量交换等宏观量)的背景下,之后的统计力学将熵表示为系统处于不同可能微观状态的概率对数函数。后者的函数形式与香农信息熵相同,其中,香农信息熵表达式中的概率是变量的不同可能取值的概率(见2.2节)。如 Edwin T Jaynes 在1957年所说那样,在更深层次上,作为用概率分布表示的不确定性度量,这两种熵的概念紧密联系;在两种情况下,都被视为受观测量约束的概率分布的预测问题,其中具有最大熵的概率分布是唯一的无偏选择[7]。
上述关于熵含义的简短论述,在 Hermann Haken 建立的协同学(synergetics)中找到一个理论框架,该框架正式集成了远离平衡系统(far-from-equilibrium systems)中耗散结构(dissipative structures)涌现的数学形式。非线性和不稳定性导致涌现和复杂性机制的产生,而熵和涨落导致不可逆性和不可预测性。这种动力学的本质自然会引出概率概念,弥补了我们无法精确刻画出系统的独特轨迹。协同学一直是将这些原则推及其它领域的驱动力,尤其是生命科学和神经科学。理清确定性和随机性影响的二元性并详述其如何在大脑中产生,将会为我们的愿景创造舞台。因此,本文的目标便是解开后面这些复杂的关系,从而统一看似无关的大脑动力学模型框架。
考虑到这些概念的互补性,且为方便来自不同背景的对宏观大脑动力学建模感兴趣的读者,我们先退一步,回顾一些概率论和信息论相关的基本概念。
>> 探索复杂神经动力学如何涌现,关注「神经动力学模型」读书会:
详情请见:神经动力学模型读书会启动:整合计算神经科学的多学科方法
Edwin T Jaynes强调,信息论的重大概念进步在于有一个明确的量,即利用概率分布来表示不确定性数量的信息熵,直观反映了宽分布比尖峰分布代表更多的不确定性,同时也满足与该直觉一致的其它所有条件[7]。在无任何信息的情况下,对应的概率分布完全无信息而且熵信息消失。在有一些确定性约束的情况下,如物理观测量均值的测量值,利用第一类拉格朗日方程(Lagrange’s theory of first kind),我们可以求解该约束下对应的最大化信息熵(等效于最小化自由能[7,12])的概率分布。在这个意义上,对熵的考量优先于对确定性影响的讨论,而且断定应符合最大熵分布,该断定基于这样一个事实,即最大熵分布符合所有确定性影响的结果,但除此之外,对其它像缺失信息这样的影响不置可否。当以熵作为主要概念时,自由能、概率分布函数和SFMs等相关量之间的关系就自然建立起来。这些关系在现实世界通过相关性表达它们自己,在物理层面上通过测量系统状态变量的函数也是可得的,而且原则上允许对系统所有参数进行系统估计。
预测编码讨论的内容及其相关概念,本质上基于三个核心方程:
第一个方程建立了贝叶斯定理的简化形式,以概率分布函数p进行表示,其中p(x,y)是状态变量x和y的联合概率,p(y|x)是给定变量x状态的条件下变量y的条件概率。在贝叶斯框架中,参数和状态变量在一定意义上具有相似地位,即它们都可以用分布进行描述并且以参数的形式进入概率函数p中。例如,给定参数k,状态x和y的联合概率可被写为p(x,y|k),确定了给定一组参数值k的条件下,获得一组数据 (x,y)的可能性,其中k的先验分布为p(k)。先验代表我们对模型和初始值的了解。
第二个方程,朗之万方程(技术上存在更精确的微积分形式可以在例如[4]中找到,此处假设是伊藤微积分)确定了生成模型,其中神经源层面的大脑活动由N维状态向量 Q=(x,y,…)∈RN 表示,f(Q,k)代表确定性影响,表示成基于状态Q和参数k(或一组参数{k})的M维流向量f。v∈RN确定了涨落影响,通常假设为 <vi(t)vj(t’)>=cδijδ(t-t’) 的高斯白噪声,其中δij是 Kronecker-delta 函数,δ(t-t’)是Dirac函数。噪声影响(包括乘性噪声或有色噪声)可能有更一般的公式,请读者自行查阅相关文献。
第三个方程确立了观测模型,通过前向模型h(Q)和测量噪声w将源活动Q(t)与实验获取的传感器信号Z(t)联系起来。对于脑电图测量,h是Maxwell方程确定的增益矩阵;对于功能磁共振成像测量,h由神经血管耦合和血流动力学的Ballon–Windkessel模型给出。在目前的情况下,观测模型无关紧要,但在现实世界的应用中非常重要,并且在模型反演和参数估计问题中通常是主要的污染因素。出于完整性的考量,我们此处提及这些工程问题,但为了简单起见,假设h是测量噪声为零的恒等运算,因此Z=Q。
预测编码涉及行为神经科学中一大研究领域[21,22],特别是致力于感知-行动和动力系统研究的生态心理学(ecological psychology)[23,24]。在这,James J Gibson 强调环境的重要性[25],尤其是有机体所处的环境如何为其提供各种行为的感知。这种感知-行动循环与预测编码中内部生成模型的预测和更新循环密切匹配。但生态心理学强调有一个细微差别,即生态学上可获得的信息——相对于外围或内部感觉——导致感知-行动动力学的出现。Scott Kelso及其同事为该框架的形式化做出巨大贡献,并开发了实验范式,用以测试生物体和环境之间协调的内部模型的动力学特性[26-28]。这些范式理论上受Hermann Haken协同学[4]的启发,而协同学一直是自组织理论的基础。这引发了大量重点关注感知和行动状态之间转换的研究工作,包括双感觉和多感觉运动协调的建模[29-32]和系统实验测试[33-44]。这些方法之后推广到更大范围的范式中[45-53],旨在提取生物体和环境的交互行为的主要特征[22]。众多工作产生的实质性证据有利于行为的动态描述,这些证据可以归入SFMs框架,用于行为[48]和大脑[54-56]中功能感知-行动变量。
我们通过计算对应的概率分布pi,从而理解决定确定性和随机性过程,该过程决定了状态变量x的离散值 xi。香农证明这样一个显著事实,即存在一个信息量 H(p1,…pn),可以唯一测度由这些概率分布所表达的不确定性[8]。他在最初的证明中指出,三大基本条件的要求,尤其是事件和概率组合定律,自然形成如下表达式。
在这里,K是一个正的常数。测度H直接对应于统计力学中熵的表达式,称为信息熵[7]。对其属性的讨论通常从观察者可获得信息量相关的详尽概率这一角度进行展开。在没有任何区别信息的情况下,拉普拉斯不充分理由原则(principle of insufficient reason)给两个事件分配同等概率。该主观思想学派认为概率是人类无知的表现,表达了我们对事件是否发生的期望。这种思想是预测编码理论及其解释大脑产生认知过程的基础。客观思想学派根植于物理学,认为概率是事件的客观属性,原则上该属性总能由随机实验中事件的频率来测度。在这,通过研究预测编码下的确定性和随机性过程,我们希望在这一点上保持中立。对确定性和随机性作用的提及通常属于客观学派的语言,其中,预测编码框架内的后续解释则是主观学派的一部分。预测编码中的生成模型通过朗之万方程表示两种类型的作用并形成概率函数,如此,通过经验测度函数<g(x)>,为我们提供了获取这一信息的途径。其中,尖括号表示期望值。
g(x)间的相关性以及归一化要求Σipi=1,表达了由确定性和随机性影响带来的约束,在这些约束下信息熵存在最大值。除使信息熵最大的赋值外,任何其它的赋值都会引入其它确定性的偏差或任意假设,而我们并未做这些假设。这种洞见是Edwin T Jaynes最大信息准则的本质[7]。因此,采用经典力学中常用的方法,引入Lagrange参数λi,我们最大化信息熵H然后得到下面公式:
其中概率分布表达式一般化为事件xi的多测度函数gi(xi)。该分布的熵S最大化,记为:
经验测度和最大信息熵之间的联系由配分函数Z确定(如状态和(Zustandssumme)中那样):
由经验测度<gj(x)>获取的相关性通过这些方程与平稳概率分布函数相联系,并且进一步联系到由朗之万方程表示的生成模型(也就是确定性影响)。下面章节中,我们将提供与神经科学明确相关的详细例子。
>> 探索大脑复杂系统的非线性与适应性,关注2024年最新「计算神经科学」读书会:
详情请见:计算神经科学读书会启动:从复杂神经动力学到类脑人工智能
虽然上述感知-行动动力学框架和预测编码互补地解释了大脑和行为如何通过作用于环境以及观察环境和自我进行塑造,但是SFMs框架给出了这些形成过程的最终结果如何体现的机制图。后者以协调的神经活动自发地涌现出流形的形式,为持续演化的内部生成模型提供了抽象相空间中时空结构的概念化,大脑便在这些流形上体验和改造世界。现在我们简要讨论在大脑动力学背景下SFMs研究中的核心动力系统概念以及相关的数学机制。
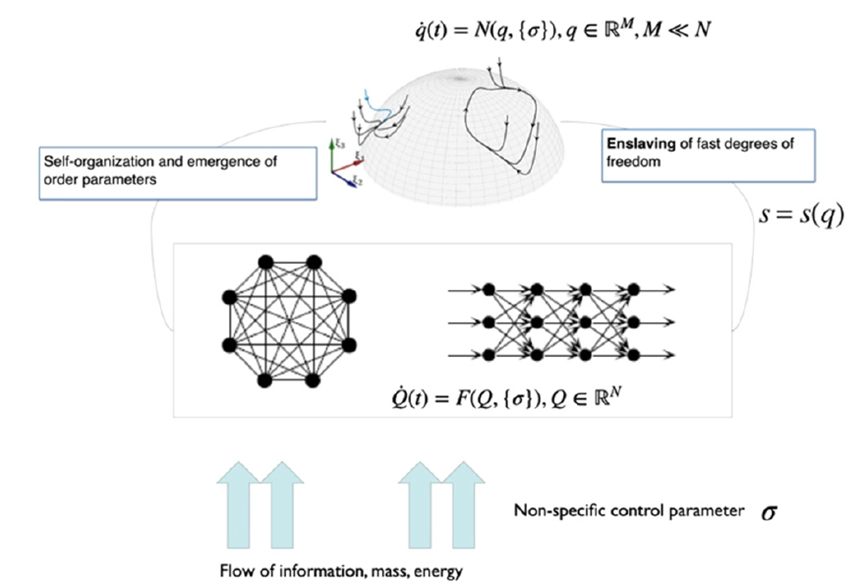
自组织系统中的涌现需要某些典型低维吸引子的稳定性出现变化。所考量的系统需是具有N个自由度的高维非线性系统。在由这些自由度构成的空间中,每个点都是一个状态向量并代表整个系统的一个潜在状态。随着时间推移,系统状态改变而因此在状态空间中形成一条轨迹。系统遵循的规则可理解为导致状态向量改变的力(作用)并定义了一个流。为了允许该系统产生低维行为,即系统维度M远远小于N,必须有一种能够将高维空间中的轨迹导向更低的M维子空间的机制。在数学上,这可以转化为两个与不同时间尺度相关的流分量:第一,低维吸引子空间包含一个流形M,其在快速时间尺度上吸引所有轨迹;第二,在流形上,结构化流F(·)规定了慢时间尺度上的动力学,在这里,慢是相对于快速动力学向吸引流形崩塌的过程而言的,见图2。为了紧凑和清晰,设想在任意给定时刻t,系统状态由N维状态向量Q(t)描述。然后将全部的状态变量分为q和s两个部分,其中q中的状态变量定义了特定任务变量M,与低维子空间(功能网络)中的涌现行为相联系,s中的N-M个变量定义了剩余的自由度。自然,N比M大得多,并且变量q子空间中的流形必须满足某些约束条件才能保持局部稳定,如此,所有的动力学才能被吸引到那里。
图2:自组织系统中的涌现。外部输入推动系统脱离由参数k控制的平衡。系统中的非线性交互导致在控制参数临界值处出现些许宏观模式(序参量)。剩余自由度被役使,并跟随着序参量演化。
首先我们概述协同学中的一些概念,这有助于更好地理解SFMs概念。协同学是远离热平衡的开放系统(即那些通过物质、能量和/或信息通量与环境相联系的系统)中自组织模式形成的理论,而这些系统由无数具有弱相互作用的微观元素组成[4]。由于微观元素之间的相互作用,这种系统可能在空间上变得有条理(有序)和时间上变成有序模式,这通常在本质上是宏观的,且可以用有限的所谓序参量(或集合变量)来描述。大脑活动的一个宏观模式失去稳定性而另一个模式占据上风时,就会发生脑活动模式的自发转换(即非平衡相变)。系统状态(或相)的稳定性是指,如果扰动使其远离,它将倾向于返回那个稳态。如果失去稳定性,系统反而会趋向于远离那个状态,而转向另一个稳态。接近这些(宏观)不稳定点时,返回的时间将大大增加。因此,宏观状态对扰动的反应演化得相当缓慢,而其底层的微观元素则保持各自的时间尺度。造成的结果是,它们动力学的时间尺度差异巨大(即时间尺度分离)。从缓慢演化的宏观状态视角来看,微观元素变化如此之快以至于它们可以即刻适应宏观变化。因此,尽管宏观模式由子系统产生,但是,打比方说,前者役使后者[4]。有序状态总是可以由少数几个变量来描述(至少在分岔附近是如此),因此,最初的高维系统的状态可以由几个甚至单个集合变量来概括,即序参量。该序参量张成工作空间。役使序参量和被役使的生成序参量的微观元素之间的循环关系有时也被称为循环因果(circular causality),这实际上允许了对系统动力学性质进行低维描述[57]。循环因果的概念和自组织系统中低维动力学的涌现是Hermann Haken协同学的核心[4]。
协同学的数学形式简述如下。N维状态向量Q(t)∈RN定义为时间函数,且包含了系统的所有状态变量。状态向量的演化由非线性常微分方程刻画:
其中,F表示刻画了所有交互作用的非线性函数,{k}表示控制参数集,控制系统的状态并且具有时间独立性。非线性演化方程包含噪声υ(t)的影响,这在之后会考虑,但非现在。状态向量及其演化方程也可进行扩展,以体现状态变量的空间依赖性,但此处仅考虑空间离散系统就已足够了。详细的数学方法见 Haken (1983)。
协同学中自组织和涌现的切入点是宏观层面系统状态的质变(状态转换)。一个给定的状态通常对应于状态空间中的一个(稳定)不动点或周期(极限)环(见图2),协同学旨在描述状态空间中其局部附近的流(见图3)。
图3:协同学和SFMs。在自组织中,涌现模式被限制在低维流形中,如球面。当流局部改变其拓扑时(左侧方形区域),协同学的临界状态出现。而SFMs的临界状态是非局域的,当整个流形的流为零时出现(临界状态)。
与稳态Q0的偏差ε(t)为Q(t)=Q0+ε(t),其时间演化可以用F在稳态Q0附近的泰勒展开式来近似:
其中L(Q0,{k})和A(Q0,{k})分别是泰勒展开式的一阶项和二阶项。随着控制参数{k}的变化,引导系统通过参数空间,遇到导致分叉的参数配置,致使当前状态的稳定性发生变化,导致系统在宏观层面上产生重大重组。分叉点定义了工作点k0,因此继状态空间工作点Q0之后,在参数空间中形成第二个局部约束。在这些局部情况下,雅可比矩阵L(Q0,{k0})的特征值λ0至少有一个变为零,允许应用局部中心流形定理。这表明,新变量的小子集Q(t)∈RM,M<<N,即序参量出现,并通过时间尺度分离的方式主导着系统的动力学,以及役使剩余变量 s(t)∈R(N-M) ,其中,序参量q(t)的动力学过程缓慢而被役使变量s(t)则迅速。在协同学术语中,役使(enslaving)是指变量s(t)的内在动力学可以被隔绝消除,其长期演化可以表示为序参量函数,即s(t)=s(q(t))。这便引出涌现动力学的简化描述:
其中P是参数为{k}和Q0的非线性函数,且可通过分解F为序参量和役使变量进行解析计算。
虽然协同学在概念上并不局限于状态空间中的局部工作点,但实际上一直如此。这极大地限制了协同框架在生物学中的使用,因为对状态空间流的探索似乎是生物体的基本活动。为此,可将对称性作为参数空间定义工作点的另一指导原则[9,16],目的是打破状态空间局部区域的限制约束。具有对称性的动力系统称为等变动力系统[59-62],群论方法提供了描述该系统的自然语言[59,60]。为证明这一点,我们考虑公式(1),且有理由用 q(t)∈RM,M<<N而非Q(t)∈RN表示状态变量。此外,为了简化,仅考虑一个控制参数k,用Γ表示作用于其解的群。如果F对于群Γ的作用是可交换的,即对于所有γ∈Γ,F(γ·q)=γ~·F(q) ,称方程为Γ等变的。对于临界值控制参数k0,这种对称性存在。Γ等变的一个重要结论是,如果q(t)是常微分方程的一个解,那么对于所有γ∈Γ,γ·q(t)也是该方程的解。如果对称性是连续的,即γ∈γ·q=q+δq,相应的群称作李群,其元素具有状态空间中M维光滑流形(记为(M))的拓扑结构,且其群运算是群元素的光滑函数。那么,方程(1)的稳态解覆盖一光滑流形M,定义为:
其允许沿流形的连续移动δq。在对称性破缺的情况下,k=k0+μ,其中μ很小,系统的解可以用全对称解(无对称性破缺时的解)的扰动来近似:
其中M为稳态解的光滑不变流形。零特征值与M的切空间相关联,这确定了与其他自由度的时间尺度层级结构,就如通常从协同学中得知那样。完全对称时,k=k0,如果M稳定,流形上的所有点都是稳定的不动点。稍有对称性破缺时,μ<<1,沿流形出现缓慢的流,这里缓慢是与流形的正交快速动力学相比而言的。这两种情况的图示见图4中的圆形流形M:0=1-X2-Y2。
图4:圆形流形上的结构化流图示。上:由于两个子系统完全对称,x,y方向上零流的两个零线重叠。圆形流形由稳定不动点组成。下:对称性破缺,如通过x和y的耦合形成,使得两个零线移开,在流形上形成一个由稳定和不稳定固定点组成的结构化流的狭窄通道。
流形上流的结构完全由N中k的对称性破缺决定。在神经科学背景中,如大规模脑网络模型,这种对称性破缺由大脑区域间连接的差异或个别区域的局部特性造成的。从传统协同学框架中我们得知,不存在导致序参量 从N维降到M维缩减的相关巨大降维因素。相反,对称性必须定义在一个由张成的子空间,然后如传统协同学框架中那样,由剩余的N-M个变量s(t)∈RN-M完成。假设这些变量在临界点k=k0处没有不稳定性,那么它们能通过通常的方式进行役使来隔绝消除,其动力学可表示为序参量q(t)的函数。传统协同学框架和SFMs之间的不同在于,前者,全N维系统考虑允许全维降维,而后者需要假设或施加额外的约束。考虑到这些,我们可以得到如下形式的系统方程:
其中
从N维降到M维缩减的相关巨大降维因素。相反,对称性必须定义在一个由张成的子空间,然后如传统协同学框架中那样,由剩余的N-M个变量s(t)∈RN-M完成。假设这些变量在临界点k=k0处没有不稳定性,那么它们能通过通常的方式进行役使来隔绝消除,其动力学可表示为序参量q(t)的函数。传统协同学框架和SFMs之间的不同在于,前者,全N维系统考虑允许全维降维,而后者需要假设或施加额外的约束。考虑到这些,我们可以得到如下形式的系统方程:
其中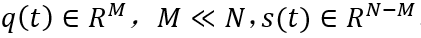 这组方程建立了由SFMs表示的基本数学框架,接下来我们将在该框架内发展出基本网络方程。
这组方程建立了由SFMs表示的基本数学框架,接下来我们将在该框架内发展出基本网络方程。
以往对SFMs涌现的讨论都是关于自然界中满足限制条件的所有动力系统的一般讨论。本节将联系与神经科相关的动力系统,旨在阐述SFMs如何自然地从基础神经科学网络中涌现以及在状态空间中创建概率分布。
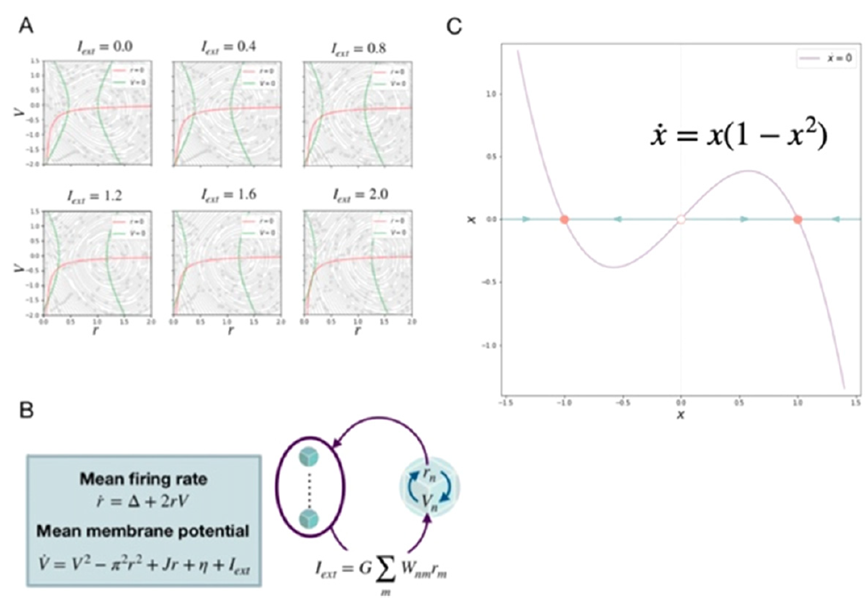
神经群体模型(neural mass model)是神经元集体活动的简化数学表示。它通常来源于由耦合点神经元模型表示的一群神经元。在对动作电位分布和/或神经元间耦合的统计假设下,应用平均场理论推导集体变量方程,刻画神经元群的均值、方差和更高统计矩的演化。显著的例子包括假设泊松分布尖峰的Brunel Wang模型[63],Zerlaut等人的使用主方程和传递函数形式的模型[64],以及利用导致神经元同步集群的神经元参数异质性进行设计的Stefanescu–Jirsa模型[65,66]。因为是在Lorentzian假设下的严密论证,Montbrio等人的平均场推导从理论角度看极具吸引力[67]。它导出了两个集体状态变量,即平均放电率r和平均神经元膜电位 V。相应的相流和方程分别见图5A和5B。这些以及其他所有神经群体模型的共同点是,将平均场动力学简化为低维表示,通常是以2维的形式。神经群体模型通常包括低放电率对应的下状态(down state),高放电率对应的上状态(upstate),以及在上状态中普遍显示振荡动力学的能力。暂时忽略振荡动力学,我们可以从概念上将共存的高、低状态,以及由分岔导致它们稳定性的改变,简化为单一变量x的相流,如图5C所示。这里将相流简化为两种稳定状态的分离,外部控制参数改变导致鞍节点分岔,可能会使系统失稳。该模型在数学上与Wong–Wang模型一致,后者是由 Brunel Wang 模型在隔绝近似的条件下推导得到。我们想要强调的是,这种表示并不是Brunel Wang 神经群体模型特有的,而是刻画了所有神经群体模型的基本动力学特性,即对于系列中间控制参数,同时存在低、高放电状态,经控制变量取高/低值的分岔,丢失低/高放电状态。因此,在这我们用该简化神经群体作为下节等变脑网络模型的基本构建块。
图5:简化神经群体模型到基本的一维形式。(A)不同输入强度的二维Montbrio模型。(B)Montbrio神经群体模型的组成。(C)简化的一维神经群体模型。
双稳态神经块组成的网络中,对称性破缺自然导致SFMs的产生[16]。这可以像之后论述那样理解。首先考虑考虑具有x,y两个节点变量的直观网络例子,其方程记为:
如此形成一个耦合神经群体模型系统,其中k为局部兴奋性,v噪声,遵循公式(1)的符号表示。图6展示了这种情况下状态空间中的相流,展示4个稳定不动点,1个不稳定不动点和4个鞍点。红色和绿色线表示零线。此时,上述方程可写成:
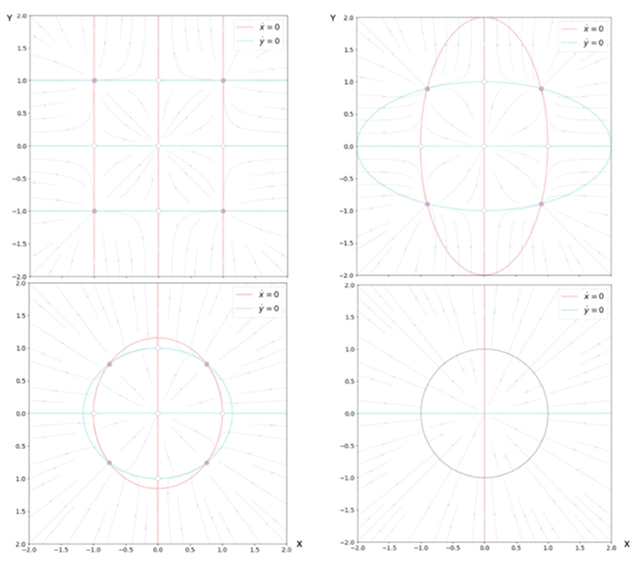
其中,第一项和前面3.3节圆形不变流形表示的一样。当G=1,这些方程与非耦合系统相同。随着G从1变为0,零线的形状不断改变,从无穷远(直线)、到椭圆再到圆形(见图6)。G作为第二控制变量,量化相互连接的程度,从完全断开(G=1)到完全耦合(G=0)拓扑。对于高度互连的网络,即G≈0,这些中间值把相流约束在闭合流形上,以原点为中心,形成对称流和时间尺度分离。如果引入其它形式的对称性破缺,如k的局部改变或不对称连接,那么这样就有一种系统地控制流形上的流的方法。
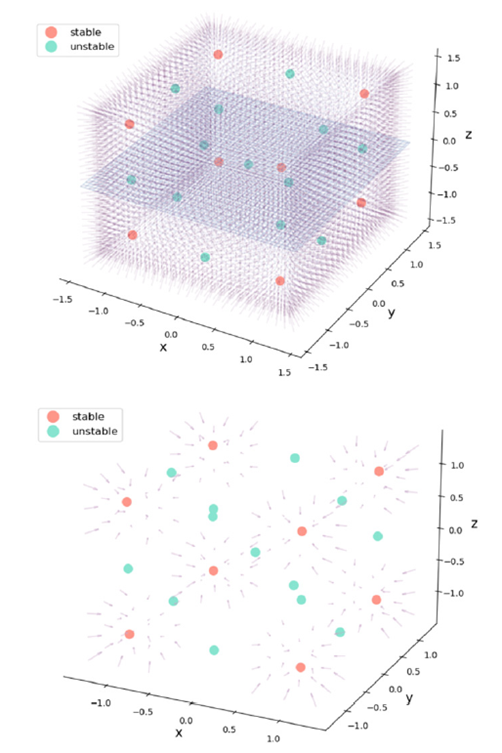
其中,G=0时在状态空间对应相流的图示见图7。此时对应于前面示例中未连接的网络节点。同样,原点是一个不稳定不动点,位于立方体中心。该立方体由1个不稳定不动点,8个稳定不动点,以及分隔这些点的12个鞍点组成,这些鞍点沿着定义立方体边缘的零线对齐(见图7)。随着三个节点之间的联通性逐渐形成,G值向着完全联通网络(G=0)方向减小。对于后者,不变流形是2维球面,以原点为中心,半径为1,且是零流。
图6:耦合参数G变化时,两个节点的状态空间流。从左上角起:G=0,0.25,0.75,1。
图7:三个耦合节点网络中的流。三维立方体覆盖一等变矩阵,中心1个不稳定不动点(绿色),8个稳定不动点(红色)和12个鞍点(绿色)。上下两图以不同的细节展示相同的情况。
该网络确定了一个 N 维状态空间中的等变矩阵,在不存在任何耦合的情况下,存在2N个稳定不动点,并由同等数量的不稳定不动点分隔,所有的不动点都以原点为中心。随着连接的建立,系统的相流越来越局限于N-1维球面 附近,也即2.2节中讨论的M流形并完全由稳定不动点组成,此时
附近,也即2.2节中讨论的M流形并完全由稳定不动点组成,此时 ,与2、3维时的情况完全类似。在更复杂的对称性破缺情况下,如连接组的引入(此时cij≠1)或区域兴奋性ki和局部噪声vi的变化,该流形上便会形成大范围的结构化流。
基于连接组的网络建模被广泛应用于静息状态的大脑活动[68-75],但是除描述性统计(如功能连接,功能连接动力学,多尺度熵)和假设机制(亚临界性,随机共振)外,还并未有严谨的理论观点。通过引入连接组,等变矩阵的对称性破缺给出一个迷人的替代性解释。但是它仍然缺少重要的解释论据,因为等变矩阵的对称性破缺并未带来实质性的降维。不变流形保持N-1维。如3.3节中论述那样,在这里特别是对于神经科学来说,问题是:N维空间中的变量Q(t)如何降维到M维空间中的变量q(t),因为M远小于N。
一个可能的实现方法如下。在更现实的神经科学模型中,不动点不会相同。通常下状态比上状态稳定。尽管这种差异不可避免地把梯度引入到网络流中,但仍不足以减少低维流形的维度,而这些低维流形是特定任务所必须的,且已知在人类大脑活动中是存在的。文献里的讨论昭示去相关作为重要信息处理机制的可能性[76-78],在这或许通过上状态中的振荡能得以实现。在动力系统理论中,利用频率分离或平均进行解耦是广为人知的一种机制(如旋转波近似),并在大规模脑网络中用于组织同步[79]和交叉频率耦合[80]。这种振荡行为的能力很适合于将等变M维流形及其N-M维互补子空间 (M<<N) 之间的连接断开,为将来的科学研究提供一条清晰的前进道路。
自由能可被视为反映了系统中存在的确定性特征产生的流形上结构化流的演化。在大规模脑网络模型的背景下,我们证明了连接组导致的对称性破缺可以作为这些特征出现的候选机制,从而与生物物理过程建立联系(如 Hebbian 学习及其它可塑性机制)。然后探索流形的随机波动以熵的形式推动系统发展。流形上的每个点都与一个概率分布相关联。没有随机力的极端情况下,这些分布具有近似 delta 函数的尖峰峰值,并且结构化流是完全确定的 [16]。这些确定和随机影响之间的联系是 Fokker–Planck 方程,它规定了概率分布函数的时间演化[4]。将我们的注意力限制在对其稳态特性的预测上,则统计特性的解释是时间无关的而且我们可以参考状态,否则系统状态在时间t时的解释将仅能在t时刻执行测量的基础上行得通。
我们希望通过两个例子来阐述这一点。第一个示例与前面4.1节讨论的情况有关,其有两个耦合节点。首先将焦点放在两个稳定不动点中的一个上。对其流进行线性化,得到如下表达式:
其中V(x,y)表示势,v表示噪声,y0表示偏移量,β表示耦合强度。确定和随机影响在实验获得的相关性中表现出来,建立在构造概率分布函数p(x,y)时需要满足的约束。函数g(x,y)的期望值是<g(x,y)>,可以由统计动量<x>,<y>,<x2>,<y2>,<xy>,…等表示。平稳概率分布函数
其中,x1=x,x2=y,N是标准化常数。根据假设可表示为:
其中所有的拉格朗日乘数λi可以通过实验获得的相关性<g(x,y)>和归一化条件
,与2、3维时的情况完全类似。在更复杂的对称性破缺情况下,如连接组的引入(此时cij≠1)或区域兴奋性ki和局部噪声vi的变化,该流形上便会形成大范围的结构化流。
基于连接组的网络建模被广泛应用于静息状态的大脑活动[68-75],但是除描述性统计(如功能连接,功能连接动力学,多尺度熵)和假设机制(亚临界性,随机共振)外,还并未有严谨的理论观点。通过引入连接组,等变矩阵的对称性破缺给出一个迷人的替代性解释。但是它仍然缺少重要的解释论据,因为等变矩阵的对称性破缺并未带来实质性的降维。不变流形保持N-1维。如3.3节中论述那样,在这里特别是对于神经科学来说,问题是:N维空间中的变量Q(t)如何降维到M维空间中的变量q(t),因为M远小于N。
一个可能的实现方法如下。在更现实的神经科学模型中,不动点不会相同。通常下状态比上状态稳定。尽管这种差异不可避免地把梯度引入到网络流中,但仍不足以减少低维流形的维度,而这些低维流形是特定任务所必须的,且已知在人类大脑活动中是存在的。文献里的讨论昭示去相关作为重要信息处理机制的可能性[76-78],在这或许通过上状态中的振荡能得以实现。在动力系统理论中,利用频率分离或平均进行解耦是广为人知的一种机制(如旋转波近似),并在大规模脑网络中用于组织同步[79]和交叉频率耦合[80]。这种振荡行为的能力很适合于将等变M维流形及其N-M维互补子空间 (M<<N) 之间的连接断开,为将来的科学研究提供一条清晰的前进道路。
自由能可被视为反映了系统中存在的确定性特征产生的流形上结构化流的演化。在大规模脑网络模型的背景下,我们证明了连接组导致的对称性破缺可以作为这些特征出现的候选机制,从而与生物物理过程建立联系(如 Hebbian 学习及其它可塑性机制)。然后探索流形的随机波动以熵的形式推动系统发展。流形上的每个点都与一个概率分布相关联。没有随机力的极端情况下,这些分布具有近似 delta 函数的尖峰峰值,并且结构化流是完全确定的 [16]。这些确定和随机影响之间的联系是 Fokker–Planck 方程,它规定了概率分布函数的时间演化[4]。将我们的注意力限制在对其稳态特性的预测上,则统计特性的解释是时间无关的而且我们可以参考状态,否则系统状态在时间t时的解释将仅能在t时刻执行测量的基础上行得通。
我们希望通过两个例子来阐述这一点。第一个示例与前面4.1节讨论的情况有关,其有两个耦合节点。首先将焦点放在两个稳定不动点中的一个上。对其流进行线性化,得到如下表达式:
其中V(x,y)表示势,v表示噪声,y0表示偏移量,β表示耦合强度。确定和随机影响在实验获得的相关性中表现出来,建立在构造概率分布函数p(x,y)时需要满足的约束。函数g(x,y)的期望值是<g(x,y)>,可以由统计动量<x>,<y>,<x2>,<y2>,<xy>,…等表示。平稳概率分布函数
其中,x1=x,x2=y,N是标准化常数。根据假设可表示为:
其中所有的拉格朗日乘数λi可以通过实验获得的相关性<g(x,y)>和归一化条件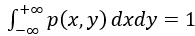 显式地估计出来。不失一般性,假设λ1=-1,λ2=0,λ3=-2β,λ4=2y0,λ5=-1,上式可简写为:
显式地估计出来。不失一般性,假设λ1=-1,λ2=0,λ3=-2β,λ4=2y0,λ5=-1,上式可简写为:
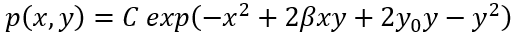
归一化因子是从具有λ0的项中获得的。这个联合概率分布函数形式上可以重写以反映贝叶斯定理同时联系到我们最初的讨论,即
其中Nx,Ny是p(x|y),p(y) 的归一化常数。如果两个节点是独立的,那么自然地动量分解<xy>=<x><y>和确定性耦合β=0。条件概率p(x|y)变成独立于y,即p(x|y)=p(x),并且p(x)和p(y)都是完全高斯的。表达式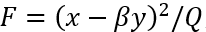 表示自由能,直观地将节点之间的相互作用刻画为静态概率密度的变形,如图 8 所示。
表示自由能,直观地将节点之间的相互作用刻画为静态概率密度的变形,如图 8 所示。
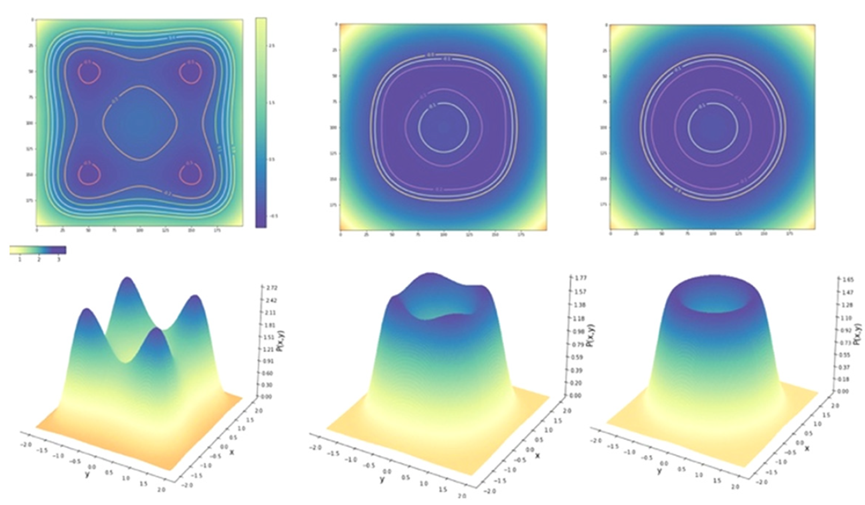
图8:耦合强度为β=1-G的二维线性耦合网络中,同一节点稳定不动点周围的概率分布。上:β=0,下:β=0.5。
另一个说明性示例如第4.2节中的等变矩阵。为简单起见,再次将讨论限制在两个节点上。按照与前一个示例相同的数学步骤,耦合节点的势函数为:
其中β=1-G表示耦合强度,β=0时表示没有耦合。同前面一样,可把平稳概率函数写为p(x,y)=p(x|y)p(y),其中
完全类比,自由能由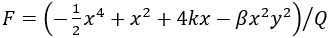 明确给出,同时给出Friston自由能原理的说明示例,其中x和y之间相关性的确定性和随机性这一基本特性通过交互作用项β来刻画。β=0,这两个节点在统计上是独立的,并且每个节点都显示出图 9 所示的双峰分布。每个单独的固定点可以用高斯局部近似,这可以通过固定点附近全概率密度的Taylor表达式解析推导出来。随着耦合强度β向1增加,平稳概率分布改变其形状,接近圆形流形并在流形上构造其结构化流。不同β值的分布如图9所示。这里的结构化流包括四个稳定和四个不稳定的不动点。
明确给出,同时给出Friston自由能原理的说明示例,其中x和y之间相关性的确定性和随机性这一基本特性通过交互作用项β来刻画。β=0,这两个节点在统计上是独立的,并且每个节点都显示出图 9 所示的双峰分布。每个单独的固定点可以用高斯局部近似,这可以通过固定点附近全概率密度的Taylor表达式解析推导出来。随着耦合强度β向1增加,平稳概率分布改变其形状,接近圆形流形并在流形上构造其结构化流。不同β值的分布如图9所示。这里的结构化流包括四个稳定和四个不稳定的不动点。
图9:双节点等变网络中的势和平稳概率分布。从左到右耦合强度为β=1-G=0,0.75,1。上面一行为势,底下一行为平稳概率分布。
如果我们接受这里讨论的熵和信息的概念作为第一个基本原语,那么根据我们对熵作为不确定性的直观理解,从信息论框架中得出的自然是以概率分布函数形状的变化表示的确定和随机影响的基本结构。这个结果适用于自然界中的所有系统,而不仅仅是大脑,这就是为什么 Hermann Haken 经常将此称为协同学的第二个基础[4]。
将视野缩小到大脑网络中存在的作用,我们将神经群体和网络的基本特性与状态空间中不变流形的出现联系起来,这是行为神经科学中,特别是生态心理学和协调动力学,已知的结构化流的载体。SFMs代表预测编码中的内部模型。这种与行为的联系很重要,因为它经常被要求指导神经科学研究,使其具有生态意义。当显式地计算概率分布函数时,自由能自然地表现为流形上的结构化流,而流形又是由网络节点之间的耦合产生的。在主动推理过程中,大脑会调整这些耦合以改变对应的SFMs(又名内部模型)。
让我们记住,一方面这些耦合(或更准确地说:耦合参数)是预测编码中自由能最小化的变化目标,另一方面负责在行为上实现特定任务的功能架构。与自由能原理处理网络在连接和参数方面“如何”(“how”)演化的机制不同,SFMs处理的是“what”——即,网络需要满足哪些约束才能实现特定流和流形的出现[16]。照此,熵和自由能可以通过学习和发展用以解释过程的演变,但也可以被视为在较短的认知表现时间尺度上发挥作用。因此,低维任务特定流形上的流在抽象状态空间中将大脑中熵的机械表现刻画为建设性的不可逆性,因此,也是神经活动和行为之间的主要使能连接。
[1] Friston K, Mattout J and Kilner J 2011 Action understanding and active inference Biol. Cybern. 104 137–60
[2] Friston K J and Kiebel S J 2009 Predictive coding under the free-energy principle Phil. Trans. R. Soc. B 364 1211–21
[3] Adams R A, Shipp S and Friston K J 2013 Predictions not commands: active inference in the motor systemBrain Struct. Funct. 218 611–43
[4] Haken H 1983 An Introduction Nonequilibrium Phase Transitions and Self-Organization in Physics, Chemistry and Biology (New York: Springer)
[5] Nicolis G and Prigogine I 1977 Self-Organization in Non-Equilibrium Systems: From Dissipative Structures to Order through Fluctuations (New York:Wiley)
[6] Varela F J, Thompson E T and Rosch E 1992 The Embodied Mind: Cognitive Science and Human Experience (Cambridge,MA: MIT Press)
[7] Jaynes E T 1957 Information theory and statisticalmechanics Phys. Rev. 106 620
[8] Shannon C E 1948 A mathematical theory of communication Bell Syst. Tech. J. 27 379–423
[9] Jirsa V K 2020 Structured flows on manifolds as guiding concepts in brain science Selbstorganisation–ein Paradigma für die Humanwissenschaften (Berlin: Springer FachmedienWiesbaden) pp 89–102
[10] Jirsa V K, McIntosh A R and Huys R 2019 Grand unified theories of the brain need better understanding of behavior: the two-tiered emergence of function Ecol. Psychol. 31 152–65
[11] McIntosh A R and Jirsa V K 2019 The hidden repertoire of brain dynamics and dysfunction Netw. Neurosci. 3 994–1008
[12] Friston K 2010 The free-energy principle: a unified brain theory? Nat. Rev. Neurosci. 11 127–38
[13] Friston K J and Stephan K E 2007 Free-energy and the brain Synthese 159 417–58
[14] Friston K J, Daunizeau J, Kilner J and Kiebel S J 2010 Action and behavior: a free-energy formulation Biol. Cybern. 102 227–60
[15] Gottwald S and Braun D A 2020 The two kinds of free energy and the Bayesian revolution PLoS Comput. Biol. 16 e1008420
[16] Pillai A S and Jirsa V K 2017 Symmetry breaking in space-time hierarchies shapes brain dynamics and behavior Neuron 94 1010–26
[17] Woodman M Mand Jirsa V K 2013 Emergent dynamics from spiking neuron networks through symmetry breaking of connectivity PLoS One 8 e64339
[18] Prigogine I 1989What is entropy? Naturwissenschaften 76 1–8
[19] Prigogine I 1987 The meaning of entropy Evolutionary Epistemology (Berlin: Springer) pp 57–73
[20] Ingvar D H 1985 ‘Memory of the future’: an essay on the temporal organization of conscious awareness Hum. Neurobiol. 4 127–36
[21] Bernstein N A 1967 The Co-ordination and Regulation of Movements (Oxford: Pergamon)
[22] Kelso J A S 1995 Dynamic Patterns: The Self-Organization of Brain and Behavior (Cambridge,MA: MIT Press)
[23] Turvey MT and Shaw R E 1999 Ecological foundations of cognition: I. Symmetry and specificity of animal-environment systems J. Conscious. Stud. 6 95–110
[24] Turvey MT, Shaw R E, Reed E S and MaceWM 1981 Ecological laws of perceiving and acting: in reply to Fodor and Pylyshyn (1981) Cognition 9 237–304
[25] Gibson J J and Carmichael L 1966 The Senses Considered as Perceptual Systems vol 2 (Boston: Houghton Mifflin) pp 44–73
[26] Kelso J S, Dumas G and Tognoli E 2013 Outline of a general theory of behavior and brain coordination Neural Netw. 37 120–31
[27] Kelso J A S, Jirsa V K and Fuchs A 1999 From level to level in brain and behavior New Developments in Statistical Mechanics ed M Bachelor and L T Wille (Singapore: World Scientific)
[28] Carson R G and Kelso J S 2004 Governing coordination: behavioural principles and neural correlates Exp. Brain Res. 154 267–74
[29] Haken H, Kelso J S and Bunz H 1985 A theoretical model of phase transitions in human hand movements Biol. Cybern. 51 347–56
[30] Beek P J and BeekWJ 1988 Tools for constructing dynamical models of rhythmic movement Hum. Mov. Sci. 7 301–42
[31] Beek P J, Peper C E and Daffertshofer A 2002Modeling rhythmic interlimb coordination: beyond the Haken–Kelso–Bunz model Brain Cognit. 48 149–65
[32] Jirsa V K and Scott Kelso J A 2005 The excitator as a minimal model for the coordination dynamics of discrete and rhythmic movement generation J. Mot. Behav. 37 35–51
[33] Kelso J A 1984 Phase transitions and critical behavior in human bimanual coordination Am. J. Physiol. Regul. Integr.Comp. Physiol. 246 R1000–4
[34] Adam J J, Paas F G, Eyssen I C, Slingerland H, Bekkering H and Drost M 1995 The control of two-element, reciprocal aiming movements: evidence for chunking Hum.Mov. Sci. 14 1–11
[35] Calvin S, Huys R and Jirsa V K 2010 Interference effects in bimanual coordination are independent of movement type J. Exp. Psychol. Hum. Percept. Perform. 36 1553
[36] Beek P J, RikkertWE and van Wieringen P C 1996 Limit cycle properties of rhythmic forearm movements J. Exp. Psychol. Hum. Percept. Perform. 22 1077
[37] Beek P J, Schmidt R C, Morris A W, SimM Y and Turvey M T 1995 Linear and nonlinear stiffness and friction in biological rhythmic movements Biol. Cybern. 73 499–507
[38] Kelso J A S 1981 On the oscillatory basis of movement Bull. Psychonomic Soc. 18 63
[39] Kelso J A S 1992 Concepts and issues in human motor behavior: coming to grips with the jargon Human Motor Behavior: An Introduction ed J A S Kelso (Hillsdale, NJ: Lawrence Erlbaum Associates) pp 21–58
[40] Kelso J A S, Delcolle J D and Schöner G 1990 Action-perception as a pattern formation process Attention and Performance XIII ed MJeannerod (Hillsdale, NJ: Erlbaum) pp 139–69
[41] Kelso J A, Southard D L and Goodman D 1979 On the nature of human interlimb coordination Science 203 1029–31
[42] Kelso J S, Southard D L and Goodman D 1979 On the coordination of two-handed movements J. Exp. Psychol. Hum. Percept. Perform. 5 229
[43] Kelso J A S and Tuller B 1984 A dynamical basis for action systems Handbook of Cognitive Neuroscience ed M S Gazzaniga (New York: Plenum) pp 321–56
[44] Kelso J A S and Zanone P G 2002 Coordination dynamics of learning and transfer across different effector systems J. Exp. Psychol. Hum. Percept. Perform. 28 776
[45] Bootsma R J, Fernandez L and Mottet D 2004 Behind Fitts’ law: kinematic patterns in goal-directed movements Int. J. Hum.-Comput. Stud. 61 811–21
[46] Huys R, Studenka B E, Zelaznik H N and Jirsa V K 2010 Distinct timing mechanisms are implicated in distinct circle drawing tasks Neurosci. Lett. 472 24–8
[47] Huys R, Fernandez L, Bootsma R J and Jirsa V K 2010 Fitts’ law is not continuous in reciprocal aiming Proc. R. Soc. B 277 1179–84
[48] Huys R, Perdikis D and Jirsa V K 2014 Functional architectures and structured flows on manifolds: a dynamical framework for motor behavior Psychol. Rev. 121 302
[49] Perdikis D, Huys R and Jirsa V 2011 Complex processes from dynamical architectures with time-scale hierarchy PLoS One 6 e16589
[50] Perdikis D, Huys R and Jirsa V K 2011 Time scale hierarchies in the functional organization of complex behaviors PLoS Comput. Biol. 7 e1002198
[51] Danion F and Jirsa V K 2010 Motor prediction at the edge of instability: alteration of grip force control during changes in bimanual coordination J. Exp. Psychol. Hum. Percept. Perform. 36 1684
[52] Fink PW, Kelso J S and Jirsa V K 2009 Perturbation-induced false starts as a test of the Jirsa–Kelso excitator model J. Mot. Behav. 41 147–57
[53] Knol H, Huys R, Sarrazin J C, Spiegler A and Jirsa V K 2017 Ebbinghaus figures that deceive the eye do not necessarily deceive the hand Sci. Rep. 7 3111
[54] GrazianoM S, Taylor C S and Moore T 2002 Complex movements evoked by microstimulation of precentral cortex Neuron 34 841–51
[55] GrazianoM S and Aflalo T N 2007 Mapping behavioral repertoire onto the cortex Neuron 56 239–51
[56] Brennan C and Proekt A 2019 A quantitative model of conserved macroscopic dynamics predicts future motor commands Elife 8 e46814
[57] Jirsa V, Friedrich R and Haken H 1995 Reconstruction of the spatio-temporal dynamics of a human magnetoencephalogram Physica D 89 100–22
[58] Guckenheimer J and Holmes P 2013 Nonlinear Oscillations, Dynamical Systems, and Bifurcations of Vector Fields vol 42 (Berlin: Springer)
[59] Ashwin P and Swift JW1992 The dynamics of n weakly coupled identical oscillators J. Nonlinear Sci. 2 69–108
[60] Chossat P and Lauterbach R 2000 Methods in Equivariant Bifurcations and Dynamical Systems vol 15 (Singapore: World Scientific)
[61] Crawford J D and Knobloch E 1991 Symmetry and symmetry-breaking bifurcations in fluid dynamics Annu. Rev. Fluid Mech. 23 341–87
[62] GolubitskyM and Stewart I 2003 The Symmetry Perspective: From Equilibrium to Chaos in Phase Space and Physical Space vol 200 (Berlin: Springer)
[63] Brunel N andWang X J 2001 Effects of neuromodulation in a cortical network model of object working memory dominated by recurrent inhibition J. Comput. Neurosci. 11 63–85
[64] Zerlaut Y, Tele´nczuk B, Deleuze C, Bal T, Ouanounou G and Destexhe A 2016 Heterogeneous firing rate response of mouse layer V pyramidal neurons in the fluctuation-driven regime J. Physiol. 594 3791–808
[65] Jirsa V K and Stefanescu R A 2011 Neural population modes capture biologically realistic large scale network dynamics Bull. Math. Biol. 73 325–43
[66] Stefanescu R A and Jirsa V K 2008 A low dimensional description of globally coupled heterogeneous neural networks of excitatory and inhibitory neurons PLoS Comput. Biol. 4 e1000219
[67] Montbri´o E, Paz´o D and Roxin A 2015 Macroscopic description for networks of spiking neurons Phys. Rev. X 5 021028
[68] Ghosh A, Rho Y, McIntosh A R, Kötter R and Jirsa V K 2008 Noise during rest enables the exploration of the brain’s dynamic repertoire PLoS Comput. Biol. 4 e1000196
[69] Deco G and Jirsa V K 2012 Ongoing cortical activity at rest: criticality, multistability, and ghost attractors J. Neurosci. 32 3366–75
[70] Hansen E C, Battaglia D, Spiegler A, Deco G and Jirsa V K 2015 Functional connectivity dynamics: modeling the switching behavior of the resting state Neuroimage 105 525–35
[71] Deco G, Jirsa V K and McIntosh A R 2013 Resting brains never rest: computational insights into potential cognitive architectures Trends Neurosci. 36 268–74
[72] Lombardo D et al 2020 Modular slowing of resting-state dynamic functional connectivity as a marker of cognitive dysfunction induced by sleep deprivation NeuroImage 222 117155
[73] Battaglia D et al 2020 Dynamic functional connectivity between order and randomness and its evolution across the human adult lifespan NeuroImage 222 117156
[74] Deco G, KringelbachM L, Jirsa V K and Ritter P 2017 The dynamics of resting fluctuations in the brain: metastability and its dynamical cortical core Sci. Rep. 7 3095
[75] GolosM, Jirsa V and Dauc´e E 2015 Multistability in large scale models of brain activity PLoS Comput. Biol. 11 e1004644
[76] Dong DW1995 Associative decorrelation dynamics: a theory of self-organization and optimization in feedback networks Advances in Neural Information Processing Systems pp 925–32
[77] Huang C G, Zhang Z D and Chacron MJ 2016 Temporal decorrelation by SK channels enables efficient neural coding and perception of natural stimuli Nat. Commun. 7 11353
[78] Daugman J G 1989 Entropy reduction and decorrelation in visual coding by oriented neural receptive fields IEEE Trans. Biomed. Eng. 36 107–14
[79] Petkoski S and Jirsa V K 2019 Transmission time delays organize the brain network synchronization Phil. Trans. R. Soc. A 377 20180132
[80] Jirsa V and Müller V 2013 Cross-frequency coupling in real and virtual brain networks Front. Comput. Neurosci. 7 78
[81] Wedemann R S and Plastino A R 2020 Nonlinear, nonequilibrium landscape approach to neural network dynamics Int. Conf. on Artificial Neural Networks (Berlin: Springer) pp 180–91
人类大脑是由数以百亿计的神经元相互连接所构成的复杂系统,吸引来自不同领域的无数科学家从不同尺度探究其中奥秘——从解析神经元之间的相互作用机理,到刻画皮层柱之间的连接形式,再到探究脑区间不同认知功能的分离与整合模式,以期揭示人脑这一最为复杂的神经系统的工作模式,理解语言、学习、社会交往等高级认知活动的底层神经机制,并有望启发通用人工智能机器人的设计。为探讨认知、智能、意识等根本性问题,集智俱乐部组织了「神经动力学模型」、「计算神经科学」、「NeuroAI」等系列主题读书会,并将在春节后开启「自由能原理与强化学习」读书会和更多活动探讨意识的量化与建模问题,探索神经科学、系统科学、生物学、物理学与人工智能如何相互启发,欢迎感兴趣的朋友报名参与。
详情请见:神经动力学模型读书会启动:整合计算神经科学的多学科方法
集智俱乐部长期深耕复杂系统与跨学科研究,在网络科学、系统理论、混沌科学等经典领域持续策划精品课程,同时也追踪因果科学、深度学习、复杂经济学、生命复杂性、社会计算、AI+Science、大模型、神经科学、城市科学等交叉前沿。汇集了大量的探索者,包括教授、硕博、工程师、创业者等等。
为了帮助有学习需求的朋友夯实学科基础,帮有科研需求的朋友深入前沿交流,我们开放了1年制和2年制的集智学园VIP计划。VIP用户可以深度参与集智俱乐部社区,并在会员期内解锁集智学园全站课程权限。