Science:统一生物异速生长的规模法则新理论
导语
生命遵循普遍的规模法则(scaling law),尤其是代谢过程,其典型是3/4次幂的异速生长律(克莱伯定律)。生物利用能量进行生长和繁衍,所以能量代谢的过程和生物的生产过程应该被紧密联系起来。在这一原则的基础上,近日发表于 Science 的文章提出了一个新理论,预测了生命的三个基本方面——代谢率、生长和繁衍之间的关系。研究发现,新陈代谢、生长和繁衍是内在联系的,它们共同决定了生物对环境的适应度。正是生物对这些因素进行共同优化的过程,导致了生物的异速生长现象,尤其是代谢异速标度关系。本文的模型还预测,人为对环境的改变将会造成物种向着降低代谢标度指数、生长率加快,以及繁衍能量(lifetime reproduction)减少的方向演化,这意味着未来生物的死亡率会系统性地增加。
研究领域:规模法则,异速生长,适应性,生活史

陶如意 | 作者
邓一雪 | 编辑

论文题目:
Metabolic scaling is the product of life-history optimization
论文链接:
https://www.science.org/doi/10.1126/science.abm7649
1. 跨越物种的统一规律
1. 跨越物种的统一规律
生物以丰富的多样性著称,但同时也存在着惊人的跨越物种的统一规律——异速生长律。
异速生长律可以描述生物内不同部位不成比例的缩放规律,如招潮蟹(Fiddler Crab)的钳子大小比它身体长度的生长要更快;同时异速生长律也能刻画不同生物之间不成比例的生长关系,如体重越大的生物,维持其生长和存活所需能量并不是成比例增长,而是和体重存在3/4幂次关系(即E=cM3/4),这也正是生物领域最耀眼的定量规律——克莱伯定律,堪称生物领域的开普勒定律[1]。
随着大数据的时代的来临,科学家们有更多数据来源可以进一步验证克莱伯定律的普适性,现如今这一结论已经扩展到了小到细胞、线粒体,大到蓝鲸将近27个数量级[2,3]。
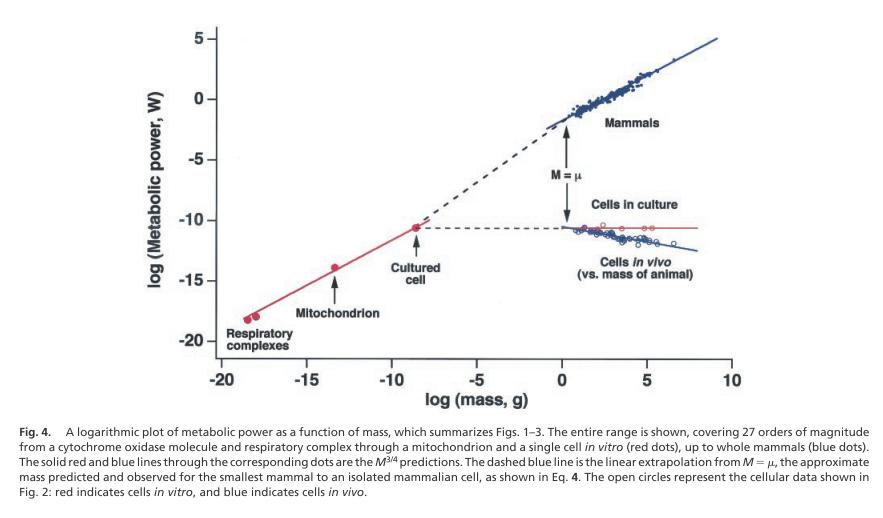
图1. 不同物种的质量和代谢率的关系。来源[3]
那么为什么生物的体重和新陈代谢会存在这样3/4的幂次关系呢?关于新陈代谢和生物的生产(生产包括生长和繁衍两个方面)之间关系的理解,存在多钟不同的理论。通常来说,有两种主要的理论,分别是新陈代谢理论(metabolic thoery)和生活史理论(life-history thoery)。
新陈代谢理论的假设是,生物代谢的废物或营养物质是通过表面积和内部网络的分布,实现物质交换或运输的,也就是说生物的几何结构约束了能量的分配模式。例如工作[4]中建模了生物的分形网络,代谢标度指数和生物的生长轨迹最终可以从这样的约束中推导出来,这是资源的获取和使用受到物理和化学约束的结果。
而生活史理论则是通过演化的视角来理解个体生长轨迹。这一理论假设分配给生存、生长和繁衍的所有资源是有限的,所以分配给每个生命阶段的资源,会受到这些阶段维持平衡的约束,这种平衡是自然选择的结果[5,6]。
本文不同于上述两种理论,为了解释个体代谢和生长轨迹,作者认为没有必要依赖约束条件。本文的思路是建模了代谢、生长、繁衍这三者的关系,而标度关系可以在自然选择过程中,最大化繁衍能量中涌现出来。
2. 代谢、生长和繁衍模型
2. 代谢、生长和繁衍模型
本文的模型建立在生物能量消耗分配的基础上。总能量支出 ET 等于生物维持自身 (self-maintenance) 的能量 EM,加上生产 (production) 的能量EP(生产能量又能分成供给生物生长 (growth) 的能量ER和繁衍(reproduction)的能量ER),以及用于消化食物的能量ED和用于活动的能量EA。所以当生物处于餐后(无消化能量)和不活动的状态(无活动能量)时,能量分配公式如下:
通常能量支出E和生物量m之间可以用幂律关系E = aE mbE来描述,aE是幂律系数,bE是幂律指数。对于总能量支出ET和代谢能量EM而言,幂指数bM往往是小于1的,而对于繁衍能量ER则是大于1的——体型越大的生物繁衍时单位体积所需要的能量是更多的,所以越大的生物繁衍过程越漫长,生的后代数量也越少。
如图1A所示,作者认为生物的生产过程在不同阶段能量的分配是不同的,在达到成熟期前,生产能量全部用来生长(EP = EG),到达成熟期后,生长能量EG逐渐减少,最终这部分能量全部分配给繁衍过程(EP = ER)。生物体质量的增加是由于生长过程提供的能量EG,即:
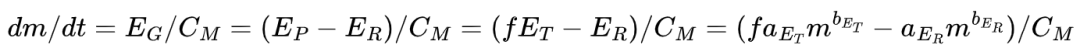
其中CM是能量和质量的转化系数,f是生产能量在总能量中占比系数。当生产能量全部分配给繁衍过程,此时生物也就停止生长,达到其质量最大值M,此时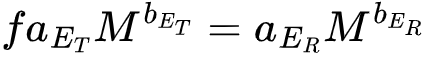 ,最终就可以估计出aER,代回上述公式就能得到生物的生长方程(图1中所示的方程)。
,最终就可以估计出aER,代回上述公式就能得到生物的生长方程(图1中所示的方程)。
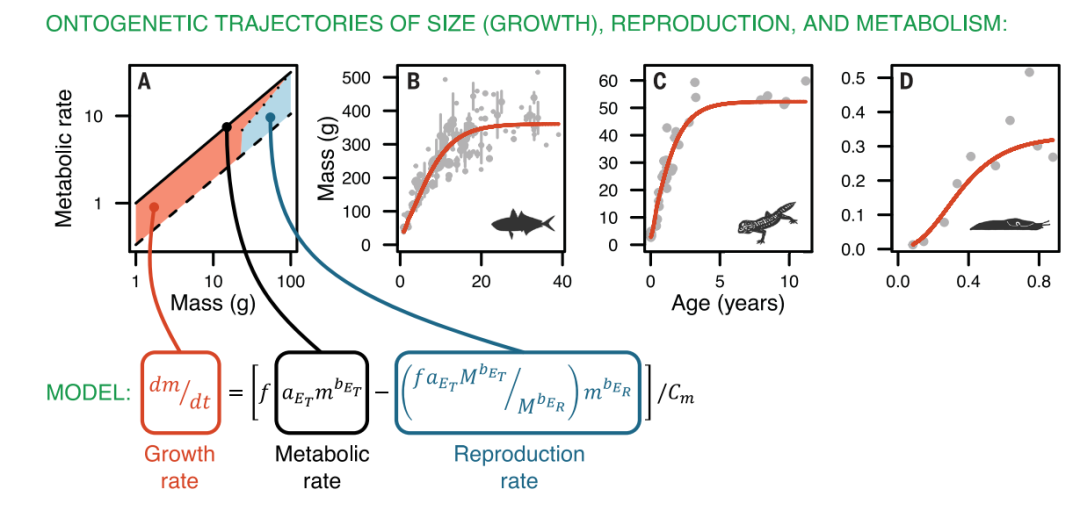
为了能够基于模型继续推导生活史的最优划分,探索代谢参数、生长和繁衍之间的关系,还有必要对模型进行一些条件的设置。这里定义了一个代谢水平(L)(可以理解为类似于每小时氧气的消耗量),相同质量的条件下,生物所需的能量ET越高会拥有更高的代谢水平L。如果这一假设成立,则意味着:
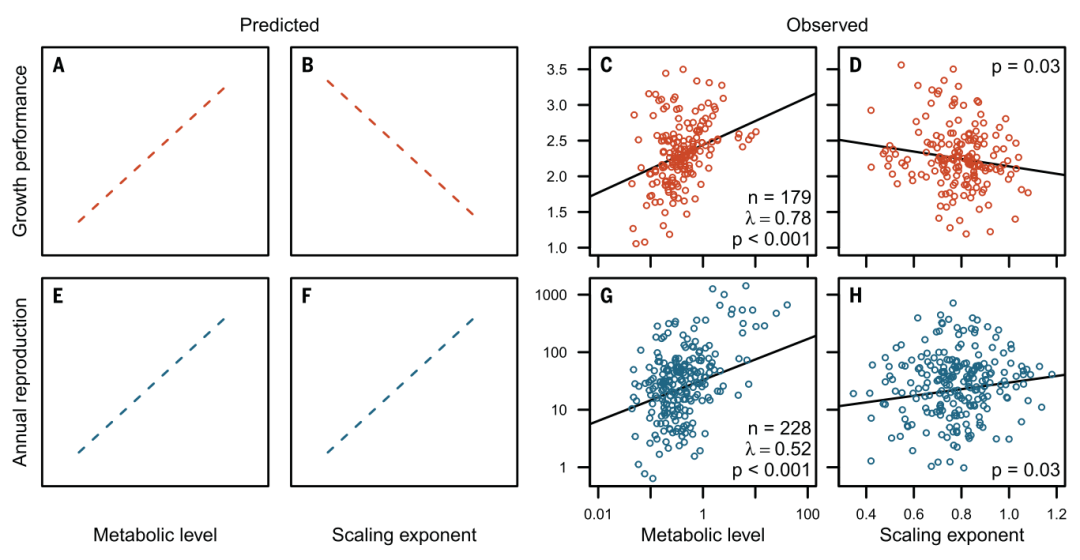
(4)给定代谢水平L,最大繁衍能量ERmax和幂指数b正相关,即新陈代谢标度更大的动物在生命后期分配更多的能量用于生产,因此有更高的最大繁衍能力(图2F)。
图2的CDGH分别对这四个预测做了验证。
3. 生活史优化
导致新陈代谢的异速标度现象
3. 生活史优化
导致新陈代谢的异速标度现象
在验证了模型的预测后,接下去作者还探索了新陈代谢水平和新陈代谢指数的影响,探究参数和最大化繁衍能量的关系。上述生长模型的繁衍能量为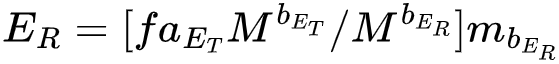 ,用数值方法估计生长和繁衍过程(其中 bER、比例 f 和成熟期质量 Mmat 的取值都参考了其他的实证工作,bER=0.137, f=0.43, m = Mmat = 223M),最终使得 ER 最大的代谢指数 bET 的值和实际观测到的代谢指数值非常接近,并且模型预测的最大 ER 会随着 bET,却不会随着代谢水平变化,也就是说用于维持代谢的能量是和物种无关的,这在7个数量级的模拟下都是如此(图3A)。
,用数值方法估计生长和繁衍过程(其中 bER、比例 f 和成熟期质量 Mmat 的取值都参考了其他的实证工作,bER=0.137, f=0.43, m = Mmat = 223M),最终使得 ER 最大的代谢指数 bET 的值和实际观测到的代谢指数值非常接近,并且模型预测的最大 ER 会随着 bET,却不会随着代谢水平变化,也就是说用于维持代谢的能量是和物种无关的,这在7个数量级的模拟下都是如此(图3A)。
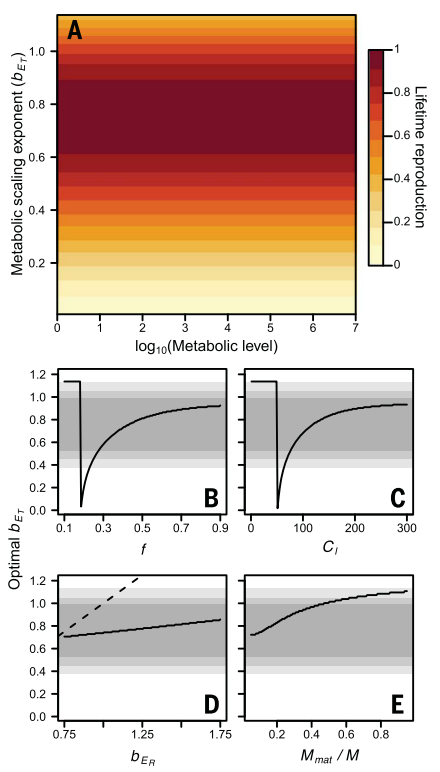
据此,作者发现了代谢的异速标度是从生长和繁衍的优化过程中演化出来的,即在有限的生命周期内最大化繁衍能量,这在某种程度上为解决生物学中最持久的争议提供了潜在的解决方案。通常来说,代谢理论解释异速标度的视角都是基于物理约束的假设,另外一些认为代谢异速标度的出现是由于自然选择的结果。本文的模型和数据其实是支持了后者的观点,认为如果自然选择在优化生长和繁衍的过程,并使得繁衍能量最大化,那么代谢理论的异速标度就可以基于此推导出来。
4. 人类世下生活史的新含义
4. 人类世下生活史的新含义
本文的框架不仅仅可以提供一个新的洞见,给出生物异速生长的起源,而且还给我们提供了一个机会去探索和预测代谢、生长和繁衍是如何应对未来环境的变化。模型预测了生物体将在未来处于增加10%死亡率的环境之中,繁衍的生命时间将持续减少,进一步将会演化为更低的代谢标度指数(从0.76降低到0.73),这和生物会增长的更快,会更早进入成熟期有关。这样的未来实在令人担忧。
本文发现展示了代谢、生长和繁衍共同演化最大化生物的适应度(如繁衍能量),并且观察到的这些生命基本特征的模式可以用优化和选择的视角——而不是约束——来解释。文章提供这一方法作为看待生物所有异速生长起源的替代方法,强调了代谢、生长和繁衍是在共演化中彼此塑造的。
参考文献
[1] Allometry. http://www.biology.arizona.edu/biomath/tutorials/applications/Allometry.html
[2] Max Kleiber. Body size and metabolism. Hilgardia. 1932, 6: 315–351
[3] Allometric scaling of metabolic rate from molecules and mitochondria to cells and mammals
[4] G. B. West, J. H. Brown, B. J. Enquist, Nature 413, 628–631 (2001).
[5] A. J. Zera, L. G. Harshman, Annu. Rev. Ecol. Syst. 32,95–126 (2001).
[6] S. C. Stearns, Funct. Ecol. 3, 259 (1989).
复杂科学最新论文

推荐阅读
-
什么是标度不变性 | 集智百科 -
社会演化动力学:人类社会复杂性为何不断增加? -
见著知微——微生物生态系统演化粗粒化模型 -
《张江·复杂科学前沿27讲》完整上线! -
成为集智VIP,解锁全站课程/读书会 -
加入集智,一起复杂!
微信扫一扫,分享到朋友圈











