最近发表于 PRL 的一项研究开发了一种新的理论框架,用于研究非平衡条件下生物化学系统中的对称性破缺。这项研究的一个重要发现是,无论生物化学反应网络的动力学细节如何,稳态下任意两组微观态之间的概率比都被限制在一定的范围内,其被称为非平衡相空间。这个范围由系统的热力学性质和网络结构决定。这个发现为我们提供了一个强大的工具来量化非平衡条件下对称性破缺的可能程度。本文是论文一作、集智俱乐部「自生成结构」读书会成员梁师翎对该研究的解读。
研究领域:对称性破缺,非平衡热力学,矩阵树定理,非平衡相空间,图灵斑图
论文题目:Thermodynamic Bounds on Symmetry Breaking in Linear and Catalytic Biochemical Systems
论文作者:梁师翎, Paolo De Los Rios, Daniel Maria Busiello
论文链接:https://journals.aps.org/prl/abstract/10.1103/PhysRevLett.132.228402
生物系统通常远离平衡态,并表现出多种选择现象。这种选择现象打破了平衡时存在的由能量确定的对称性。例如,在DNA复制过程中,生物体能够以高保真度选择正确的核苷酸,从而确保遗传信息的准确传递,保证生物的生存。这种选择机制的准确性受到平衡态下能量的限制,但生物体可以通过消耗能量的中间步骤来提高选择效率,从而实现更高的复制保真度。
此外,空间上的对称性也可能被反应扩散过程打破,Turing在1952年提出的开创性工作中阐述了这一点,其发现在某些条件下,反应扩散系统中的物质均匀分布的状态会变得不稳定,从而出现空间斑图,例如动物的毛皮上的花纹 [1]。
自从Prigogine等人提出耗散结构的概念以来,热力学与对称性破缺的关系一直是人们理解自然界的重要课题 [2]。然而,由于非平衡系统动力学的复杂性,我们一直缺乏普适的方法来理解对称性破缺的普适热力学原理。
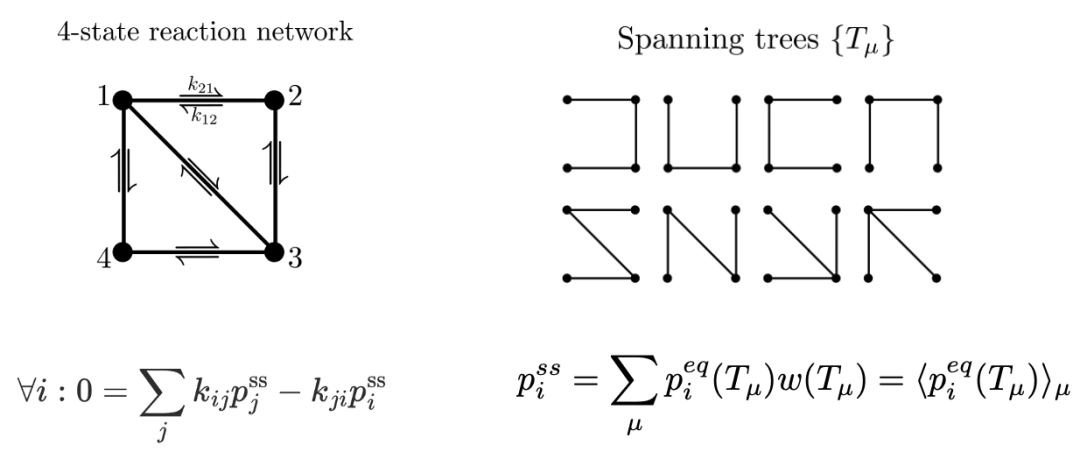
在最近发表在 PRL 上的一项研究中,来自瑞士洛桑联邦理工学院(EPFL)和德国马克斯普朗克复杂系统物理研究所的研究人员开发了一种新的理论框架,用于研究非平衡条件下生物化学系统中的对称性破缺 [3]。他们利用矩阵树定理(matrix-tree theorem, 一种将图论应用于线性代数的定理),将稳态的概率与生成树(spanning trees)的概念巧妙地联系起来,推导出了由热力学确定的稳态下任意两组微观态之间概率比值的上限和下限,并发现这些边界仅取决于网络几何形状和热力学性质,与具体的反应动力学无关。
图1. (a) 一个有四个状态的化学反应网络, 其稳态解由主方程给出。(b) 矩阵树定理允许我们将非平衡稳态解表示为系统所有可能生成树(spanning trees,即包含网络中所有节点的树)上的平衡态解的加权平均。
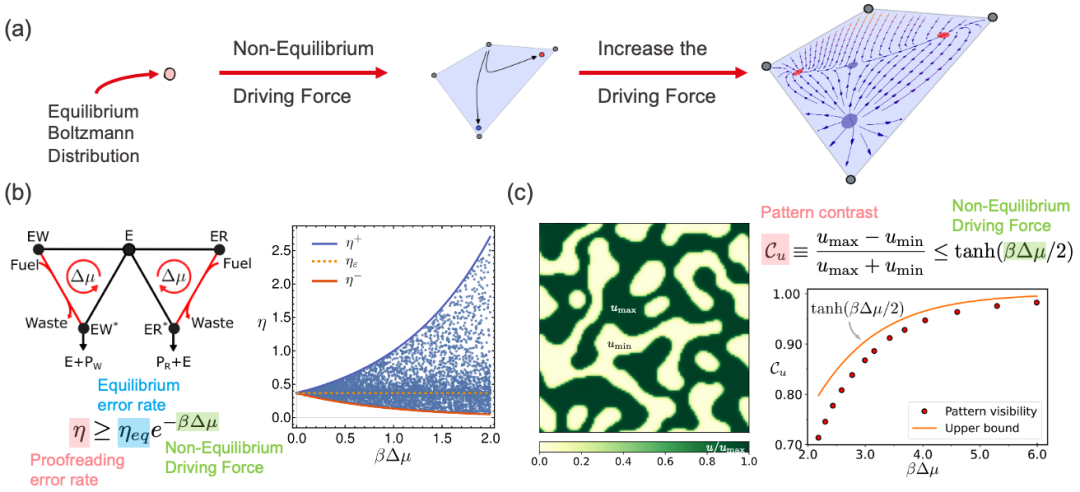
这项研究的重要成果之一是,发现了稳态概率比值的热力学上限和下限。这意味着,无论生物化学反应网络的动力学细节如何,稳态下任意两组微观态之间的概率比都被限制在一定的范围内,其被称为非平衡相空间。这个范围由系统的热力学性质和网络结构决定。这个发现为我们提供了一个强大的工具来量化非平衡条件下对称性破缺的可能程度。非平衡化学网络的复杂动力学被这个非平衡相空间所限制住,在其之中,各种生物功能得以实现,比如通过动态校对降低错误率,通过图灵模式形成斑图。
图 2. (a) 非平衡驱动力能打开原本在平衡态下无法达到的化学空间,即非平衡相空间,其表示系统所有可能的非平衡稳态解。有限的非平衡相空间允许复杂的动力学机制产生。(b,c) 非平衡相空间的概念帮助我们理解生命过程中的普遍热力学限制,比如动力学校对过程的错误率,反应扩散斑图的对比度,都受到非平衡驱动力的限制。
该研究可用于研究动力学校对和反应扩散斑图,并还可应用于化学主方程(化学反应网络的随机动力学模型)以及各种可能的生物化学功能。除了对非平衡稳态的热力学限制外,对时间反演对称性破缺的基本热力学限制在生物中也具有重要意义 [4]。
此工作的结果可对生物学、化学和物理学等多个领域产生广泛影响,例如,在理解生命起源问题上,手性对称性破缺的出现(即手性分子与其镜像分子的不对称合成)是一个有趣但尚未得到彻底探索的开放问题,这项研究的结果可能有助于理解非平衡条件在其中的作用。此外,这项研究还可能为设计人工生命系统和生物分子机器提供新的思路。
[1] Alan Mathison Turing. “The chemical basis of morphogenesis”, Phil. Trans. R. Soc. Lond. B23737–72 (1952).
[2] G. Nicolis and I. Prigogine: Self-Organization in Nonequilibrium Systems. From Dissipative Structures to Order through Fluctuations. J. Wiley & Sons, New York, London, Sydney, Toronto (1977).
[3] Liang, Shiling, Paolo De Los Rios, and Daniel Maria Busiello. “Thermodynamic Bounds on Symmetry Breaking in Linear and Catalytic Biochemical Systems.” Phys. Rev. Lett. 132, 228402 (2024).
[4] Liang, Shiling, and Simone Pigolotti. “Thermodynamic bounds on time-reversal asymmetry.” Phys. Rev. E 108, L062101 (2023).
生命是有心(意识)的,但当你一层层剥开生命的组织,却只有心脏而没有一颗“心灵”;打开大脑皮层看到一个个神经元,却看不到“意识”。然而,生命与意识都具有“自我生成“的能力,生命自发从非生命中生成,意识自发从生命中生成。更惊人的是,生命与意识的自我生成结构似乎很相似。如果这个假设成立,那么最可能的备选结构会是什么呢?
为了更深入地认识复杂活系统的自生成结构,集智科学家小木球(仇玮祎)联合周理乾、王东、董达、刘宇、苑明理、傅渥成、章彦博等科学哲学、计算机科学、物理学和生物化学等学科的一线研究者共同发起组织《自生成结构系列读书会》。其中第一季《共识——自生成结构与自复制自动机的研究背景》已经完结,读书会所形成的自生成结构社群集结了相关领域的教授、硕博及产业界人士。现在报名读书会即可解锁第一季读书会所有录播视频并入群,欢迎从事相关研究、对相关领域有浓厚兴趣的探索者报名参与交流。
点击“阅读原文”,报名读书会