生命是什么?圣塔菲研究所提出“个体信息理论”,重新定义个体生命

个体之于群体,如滴水之于百川。但在对生命本质探索中,科学上个体的概念一直模糊不清,既没有通用定义,也没有明确划分标准。最近圣塔菲研究所现任所长 David Krakauer 发表了一篇文章,基于过程哲学和演化格式塔方法,他认为个体是一种能将自身信息从过去传播到未来、并保持一定时间完整性度量的集合体,并用信息论给出了概念的形式化定义。从公式中可以推导出原则上不同的三种个体:有机个体、群落个体和环境驱动个体,每种在环境依赖和信息继承程度方面都有所不同。即使没有体细胞等明确物理边界,在合适的观察者和测量下,从分子到文化,一些多层次、高分布的适应性系统,也可以被识别成一个生命个体。文章关于个体和环境关系的刻画也为定量研究生命提供了新的理论基础。
一、从生命到个体:信息论的建构
生命本身让人充满惊异,无数个生命更是如此。
在生物学上,人类和大猩猩、海豚、狮子、蜜蜂、蚂蚁等一样都是社会性动物,群居利他,不能独存,存在极其复杂的群体行为模式。在有着庞大而悠久历史的种群面前,个体生命似乎微不足道,甚至人们一谈起生命,都默认至少是一个物种,或者干脆超越所有物种之上的某个整体。

但有趣的是,同样作为生命,一种社会化动物,尽管对人类是否是具有像蜜蜂和蚂蚁那样高度的社会性还有争议,但似乎从来没有哪一种动物能像人类那样注重个体价值。不管公权力怎么诠释,追求个体自由和自我实现都已成为这个具有共识想象力的物种越来越普世价值的一部分。对很多现代科学家,如动物行为学家和社会心理学家而言,人类就是一种“合作的物种”,在深处甚至会牵扯到复杂的演化博弈论和经济学研究视角解读。
其实远不止在生物学和心理学层面。之于社会学和政治学,对人类本性的追问,也往往在个体和群体二者之间展开。亚里士多德早就说过“人是城邦的动物”。先有个体(individuality),还是先有社会?这一历史之问,让如今人类社会政治理念之争的焦点,更是在自由主义和社群主义之间激烈而持久展开,从政治、经济、文化到生态全方位影响着整个人类发展前景。
可以看到,尽管个体与群体、甚至生命的关系如此重要,但关乎个体最根本一些问题依然还没有被解答,甚至根本没有认真人追问这个问题:
——究竟是什么才是个体?
对这个问题的回答不仅关乎个体,关乎人类整体,更是关乎生命意义本身。
在生物学中通常存在的个体性假设,就像经典时空观一样,往往只是基于天然的生物学特征,不仅没有共识,也没有严格的量化方法来识别。
最近,圣塔菲研究所现任所长 David Krakauer 在Theory in Biosciences期刊领衔发表了一篇文章,对个体本质进行了回答。他认为个体是一种能将自身信息从过去传播到未来,并保持一定时间完整性度量的集合体,并用信息论和图示给出了概念的形式化定义。个体信息理论从信息熵和互信息公式中可以推导出原则上不同的三种独特形式个体:有机个体、群落个体和环境驱动个体,每一种在环境依赖程度和信息继承方面都有所不同。
这种新型的进化格式塔方法允许扩大生命的定义范围,即使没有细胞壁或体细胞等物理边界组织,在合适的观察者和测量原理下,从分子到文化的所有组织层次,一些可以通过粗粒化减少不确定性的多层级、多尺度、高分布适应性系统,也可以被看作是一个生命个体。
论文题目:
The information theory of individuality
论文地址:
https://link.springer.com/article/10.1007/s12064-020-00313-7#Sec10



信息的起源

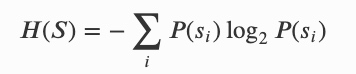
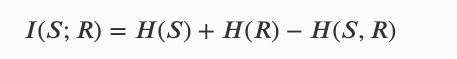
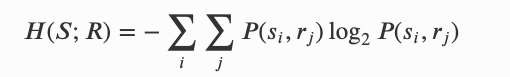
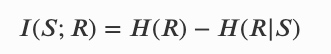
![]()


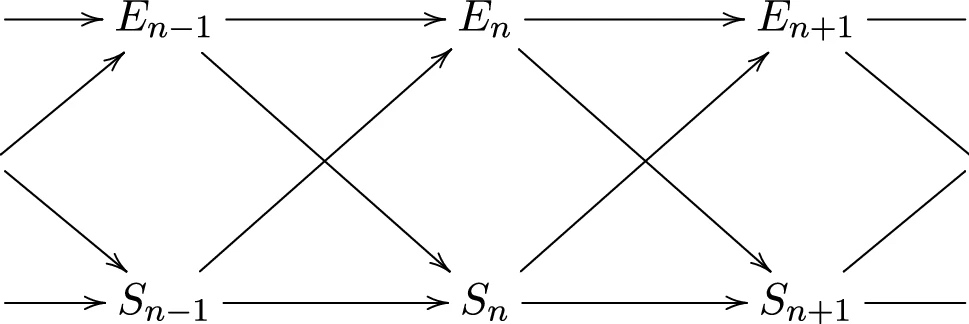
图 6:系统环境作用因果图
信息型个体
-
系统-环境分解:类比于格式塔心理学的图形-背景关系,一个动态的可测集合,可以粗略划分为个体系统和环境两部分(主体-环境)。这种划分可容许各种系统层次存在,如自然和生物上细胞器、细胞体,乃至生物体、种群、文化。 -
信息个体:一个一般离散的随机过程,未来状态由当前状态的子集决定的。当所有状态分为个体和环境两部分、并分别具有状态 𝑆𝑛 和 𝐸𝑛 时,就足以预测下一个个体状态 𝑆𝑛+1,形式上可以用互信息来量化:
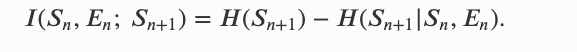
这个公式表达在时间 𝑛+1 时,个体 𝑆𝑛+1 有多少信息来上一个时间段的系统自身(前代)𝑆𝑛 和 环境信息 𝐸𝑛。其中程的互信息可以有两种方式分解: 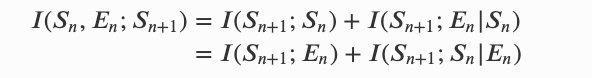
-
第一项 𝐼(𝑆𝑛+1;𝑆𝑛) + 𝐼(𝑆𝑛+1;𝐸𝑛|𝑆𝑛) 是系统对自身状态影响(下代或下一时间)的一种度量,在所存在时间间隔中,连续系统状态之间所有观察到的依赖关系都归于系统自身。Krakauer 和 Zanotto (2006) 将量 𝐼(𝑆𝑛+1;𝑆𝑛) 称 为自主性(autonomy),下面用 𝐴∗= 𝐼(𝑆𝑛+1;𝑆𝑛) 表示。当个体能控制自身时,它应该是高的。
-
𝐼(𝑆𝑛+1;𝐸𝑛|𝑆𝑛) 则可以理解从环境流向个体的新信息。若无则说明一个系统是信息封闭的,因此这个量衡量了个体受环境控制的程度, 用 nC = 𝐼(𝑆𝑛+1;𝐸𝑛|𝑆𝑛) 表示。注意,封闭性并不要求因果独立,只是说明所有来自环境的影响都是个体可以预测的。
-
群落个体 𝐴 := 𝐼(𝑆𝑛+1;𝑆𝑛|𝐸𝑛)(Colonial Individuality)
-
有机个体 𝐴∗ :=𝐼(𝑆𝑛+1;𝑆𝑛)(Organismal Individuality)
-
环境驱动个体 nC := 𝐼(𝑆𝑛+1;𝐸𝑛|𝑆𝑛) (Environmental Determined Individuality)
在使用互信息的链式法则,将影响归因于环境或系统时会遇到了一个模糊的问题:部分信息分解问题(Williams & Beer, 2010; Bertschinger et al., 2013)。通过引入独有信息、共享信息和交互信息的概念可以解决这点[1]。
在时间 n+1 的未来个体状态与时间 n 个体与环境的联合状态之间的互信息,可以可以被分解为四项:

这四项出现在前面从链式法则中得到成对的互信息和条件互信息中:
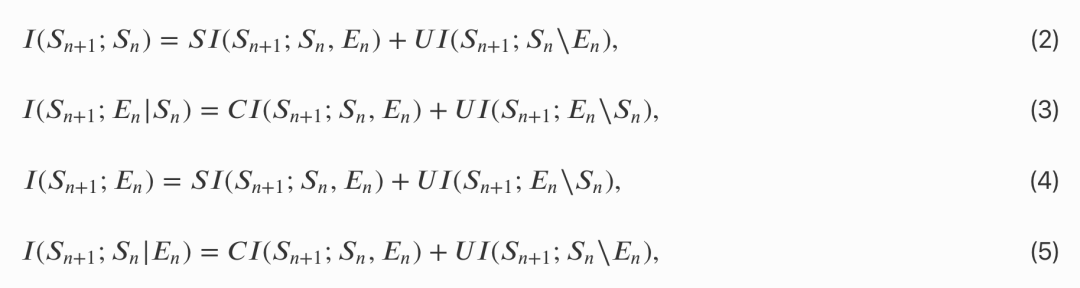
在本文研究定义的语境中,这四个词分别含义如下:
-
a 个体信息 𝑈𝐼(𝑆𝑛+1;𝑆𝑛∖𝐸𝑛):只来自个体自身并由系统自身维持; -
b 共享信息 𝑆𝐼(𝑆𝑛+1,𝑆𝑛,𝐸𝑛):个体和环境之间共享的信息; -
c 环境信息 𝑈𝐼(𝑆𝑛+1;𝐸𝑛∖𝑆𝑛):环境对个体影响的信息,狭义上即感官的信息流; -
d 交互信息 𝐶𝐼(𝑆𝑛+1;𝑆𝑛,𝐸𝑛):协同信息,只存于个体系统和环境的交互作用中。
这些分解只是作者视角下一种形式,在 Journal Entropy 中可以看到其他替代性见解(Lizier et al., 2018)。但基于此,接下来就可以严格定义三种不同形式的个体了,并且能量化每种信息的对不同类型个体的独特贡献。
例如,系统和环境共享的信息(如自适应信息)、个体或环境独有的信息(二者不同的的存储器)以及以某种复杂方式依赖于二者交互作用的信息(如控制信息)。
有机个体 𝐴∗
𝐴∗ = 𝑆𝐼(𝑆𝑛+1;𝑆𝑛,𝐸𝑛) + 𝑈𝐼(𝑆𝑛+1;𝑆𝑛∖𝐸𝑛)
当生物体能通过适应或学习与所处的环境共享重要信息时,它们就具有良好的适应性。此外有机个体还包含了有效运作时所需要的大量私有信息。通过最大化这种度量,我们能够在环境中识别出复杂的有机个体。
群落个体 A
𝐴 = 𝐶𝐼(𝑆𝑛+1;𝑆𝑛,𝐸𝑛) + 𝑈𝐼(𝑆𝑛+1;𝑆𝑛∖𝐸𝑛)
环境驱动个体 nC
环境编码
为了更好理解这些个体度量,原文举了一个可视化例子进行量化研究。
遵循前面个体形式信息定义和图 6 中介绍的一般结构,考虑两个初始状态为 {-1,+1} 的二进制集合 𝑆𝑛 和 𝐸𝑛,其中状态按照指定的条件条件分布同步进行更新。系统状态 𝑆𝑛+1 初始条件包括 𝛼𝑆、 𝛼𝑆、 𝛾𝑆 三个耦合参数,环境参数状态则是 𝛼𝐸、𝛽𝐸、𝛾𝐸。
-
𝛼𝑆:个体系统状态与自身其前一状态 𝑆𝑛 的耦合参数,如生物各种内部调节系统。 -
𝛽𝑆:个体与环境交互作用的耦合参数。如生物的自适应性。 -
𝛾𝑆:调节原个体系统与环境状态综合影响的参数。是一种高阶相关性。如并非直接和生物交互作用的自然选择或各种偶然因素。
把每一种个体度量标准都用到这个随机过程上。对随机环境分析结果如图 7 所示,对有记忆环境结果如图 9 所示。
随机环境下三种个体
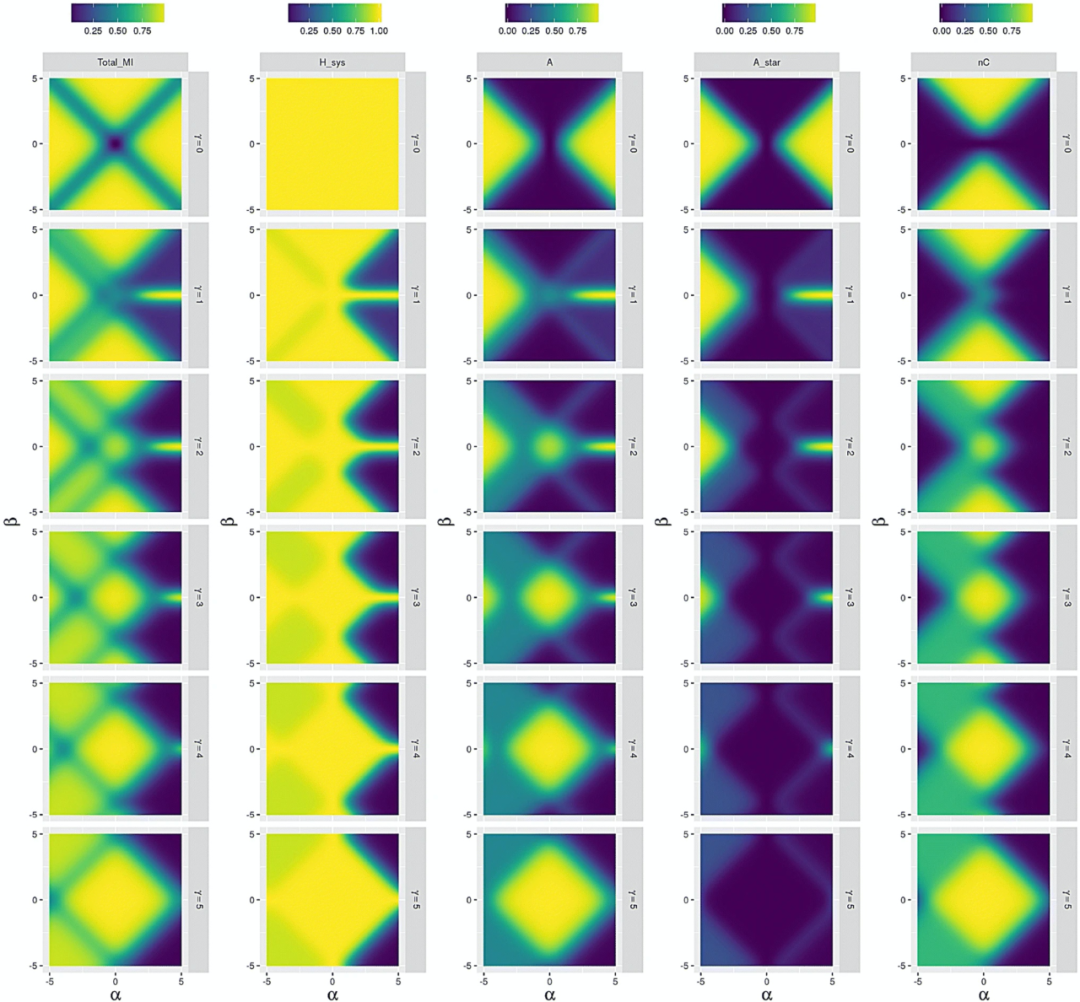
图 7:随机环境 𝛼𝐸 = 𝛽𝐸= 𝛾𝐸 = 0 下,系统互信息(Total_MI),系统熵(H_sys),群落个体 (A) ,有机个体(A_star)和环境驱动个体 (nC))对 𝛼𝑆,𝛽𝑆 和 𝛾𝑆 图示,下标 𝑆 在图中已省略。其中的系统信息值指针:越偏向蓝色越低,越偏向黄色越高
当 𝛾𝑆=0 时,个体和环境时间序列之间没有任何额外高阶相关性(如个体不能改变环境)。这时当 𝛼𝑆 和 𝛽𝑆 较高时,理论上最容易检测到群落个体和有机个体。当也正因为和环境没有高阶相互作用,这两个参数就都代表了系统状态唯一信息,因此两种个体类型此时变得不可区分。只有随着更多信息不断传递到未来,它们之间的区分才会变得更明显。
例如,最近海洋生物学家们在澳大利亚西海岸的 Ningaloo 地区出海考察,发现一只长达 47 米的巨大水母(Apolemia)[3]。但它其实不是一个简单的独立的有机个体,而是由数百万个无性繁殖的个虫(zooids)互相连接组成的巨型生物群落,每个部分都负责不同的工作,有的负责捕食、就的负责消化、还有其它的个体掌管感知、运动、繁殖等等不同功能。

图 8:长达 47 米的水母群
显然,如果这一生物存于与外部没有交互的环境中,例如在博物馆或小说中,我们是很难分清它究竟是一个有机个体,还是一个群落个体的。但以目前标准判断,它是一只典型的群落个体。
有记忆环境下的三种个体
所谓非随机有相关性的环境,即有规则和记忆的环境下,个体和环境存在着持续交互作用。
在图 3 中,环境耦合参数 𝛼𝐸 =2, 𝛽𝐸= 𝛾𝐸 = 0,同样展示了互信息、信息熵,与群落、有机和环境驱动三种不同个体在不同 𝛼𝑆、 𝛽𝑆、𝛾𝑆 下的状态图示。
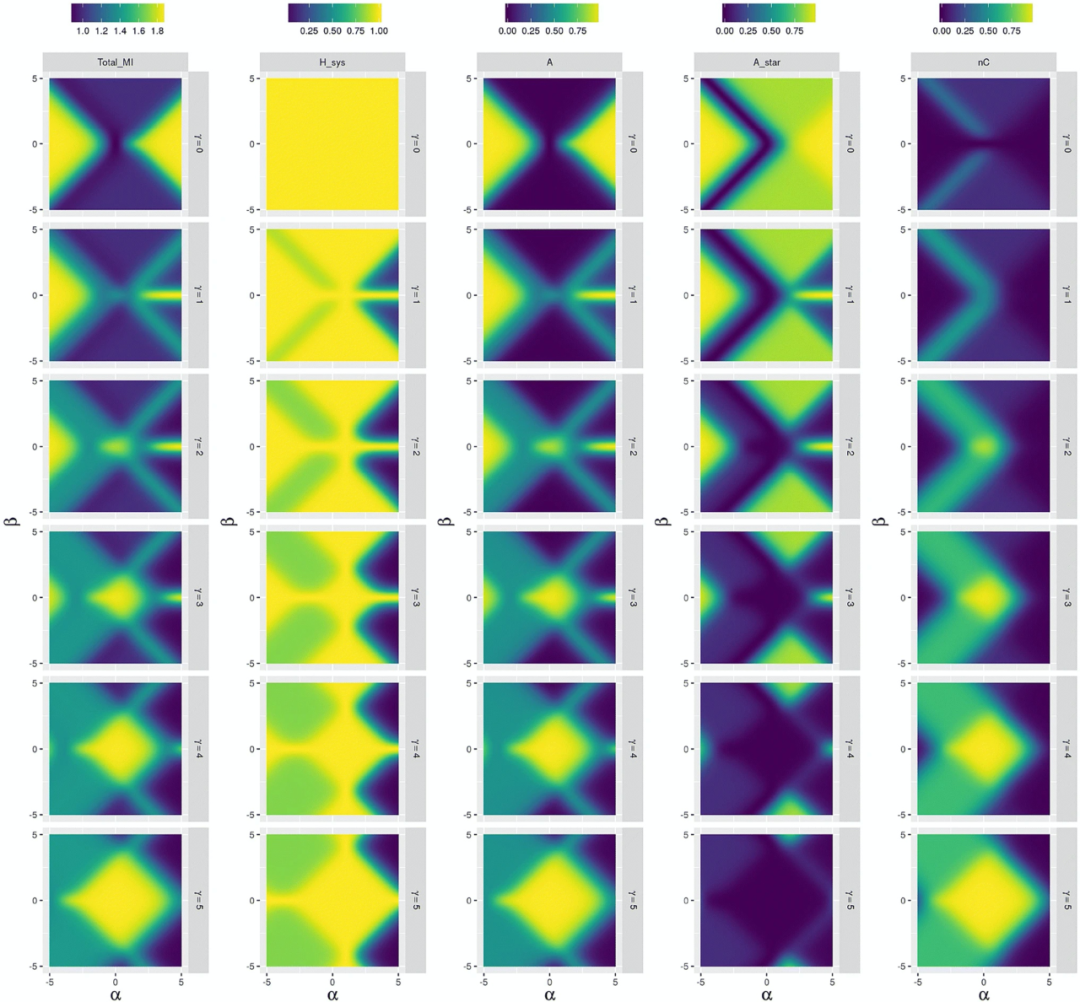
因为此时个体能够适应环境,在较高的 |𝛽𝑆| 和较低的 𝛾𝑆 下,环境不因为生物改变但会持续进行交互作用的情况下,即可以观察到较高值的有机个体 A*,以及较低值的群落个体 A 和环境驱动个体 nC。(换句话说,正因为存在交互,是在随机环境情况下由高 nC 值代表的环境中信息流到了个体系统、并被内化到个体系统中去了。)
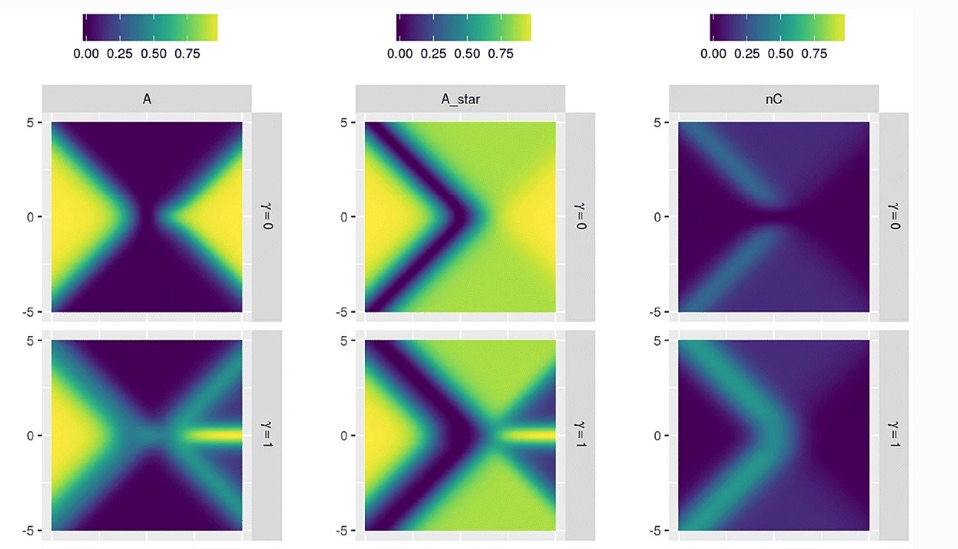
图 10:(图 9 局部)低 𝛾𝑆 下,具有较高值的有机个体 A*,较低值的群落个体 A 和环境驱动个体 nC
但随着 𝛾𝑆 值增加,有机个体和群落个体特征发生了分歧。
在低值 𝛼𝑆 和低值 𝛽𝑆 时,群落个体最显著,大部分持久性信息来自于系统和环境之间的持续互动。大部分具有社会化属性的动物,即使很弱社会性的独居动物,只要是具备最小两性生殖互动的生物,都属于这种情况。
而一般的有机个体则在高 𝛾𝑆 时,因为自主性丧失信息开始消失。只有在只有在高 𝛼 的情况下,也就是说环境对它几乎没有影响的情况下、只和自身相关的情况下,才会被保留下来。这不由得有让人想起能在极为苛刻条件如-272℃低温和151℃高温、数千米海拔和很低、甚至外太空下能生存的水熊虫(Water Bear),生物学家一直很好奇它究竟有什么生理机制、靠自己就能活的如此顽强。
图 11:被誉为地表最强生物的水熊虫
至于环境驱动个体 nC 的信息,在低 𝛾 时转变为群落个体,到高值 𝛾𝑆 时几乎难以区分。这是因为,当个体和环境变得强耦合时,交互信息(complementary)将占据主导,环境本身对个体未来状态的预测性就会降低。
显然,𝛾𝑆 的作用是通过在信息通道中创建系统的相关性和规律性,降低系统的总熵(H_sys),并在时间序列中持续反转总互信息的模式。
在低 𝛼𝑆 和低 𝛽𝑆 时,H_sys 分别在 𝛾𝑆=0 和 𝛾𝑆=5 时取得最小值和最大值。
从前面实证例子中,可以发现一个在更普遍在环境中识别不同形式信息个体的过程。当高阶耦合度(higher-order coupling)较低时,个体-环境的区别随着独立记忆的参数(𝛼,𝛽)而增加(即 𝛼 和 𝛽 增加分别意味着个体系统自我维持,和与环境进行交互作用的程度增加)。
然而,当高阶耦合度参数持续增加时,有机个体趋于消失,群落个体随着独立记忆减少而出现。
这就意味着,当生物与环境存在高阶相关性的时,物种为了在更广阔环境下具有更高的适应性,成为群落个体将是一个必然趋势。这一趋势会伴随着有机个体特性的消失,包括自身独立记忆信息的减少。
显然,我们在蜜蜂和蚂蚁这些具有真社会性的生物中,就看到了这种趋势。它们从一生下来就作为集体的一部分而存活,被划定好了分工,不用对比人类,即使相对大猩猩或老虎这样群居甚至独居动物,单独个体就几乎已经失去了自身记忆和生存能力。

图 12:作为真社会性生物整体的蚁群,个体几乎没有意义,在协调整体和路径计算上却具有很高的智能
选择的要求和层次
Krakauer 这篇论文将自适应个体的讨论建立在了坚实的逻辑和概率基础之上,为此囊括了任意粒度尺度和有意义时间长度能力。但同时因忽略了如生物细胞膜之类的特定层级功能,默认生物个体是在无偏假设下长期存在、具有复制能力的聚合体,作为选择的运作的个体层次。这符合大多数人观念中生物个体性的隐含(Okasha, 2006)假设。
然而,显然在不同时空尺度、标准和选择层次下,作为信息个体的界限将是极大不同的。
为此论文作者讨论了识别前个体(低自主性)和完全成熟个体(高自主性)的一些原则和需要进一步研究的工作:
-
划分性要求(The partitioning requirement)
例如在一篇科幻小说中,天外来物如石头一般一动不动,人类甚至对其进行了敲击,也没有丝毫反应。但过了上千年,它竟然开始活动了。原来是它只是反应时间极长的一种生命体。可见时间长度选择的重要 。在非常短或非常长的时间尺度上,不太可能观察到个体特性的量化规律,包括自主性( autonomy)、封闭性(closure)和充分性(sufficiency)等。在空间尺度的选择也类似,例如在盖亚假说中,就认为地球本身就是一个巨大的有机生物体。甚至有人认为宇宙本身,或者基本粒子都可能具备生命某些特性。
-
鲁棒性要求(The robustness requirement)
鲁棒性要求除了确定系统嵌套或层级之外,还需要对产生时间序列的生成器机制本身进行一定的规范。这等同于对个体具备一定程度的鲁棒性或纠错特性的要求。毕竟个体的自适应系统中与自适应平衡机制,即监测内部状态并确保偏差最小化有关。正是个体这种自我保护特性,能让人们在不夸大动态差异的情况下对物理和生物现象作出一些有用的区分。
-
选择的层次(The levels of selection)
在以往许多论述中,已有人提出个体具有特殊的进化地位的观点:即在一定选择层级上,较粗粒的聚合体与环境协调具有了持久性属性后,再将个体视为了独立的选择性单元。
对这个过程最流行的形式化方法是演化生物学中的普莱斯方程(Price Equation):该方程描述了一个特征的平均值是如何随着与之前性状的值、以与及适应度的协方差而变化的。它将性状特征的值分成若干个组,并试图以最好的方式来描述性状均值在种群中的演变。对此,如果确实存在一些演化的底层结构和动力学过程(见附录, Nowak et al., 2010),该方程的准确性将取决于对分类的选择(Krakauer & Flack, 2010b),而 ITI 恰好可以为建模提供这样一个基础性的平台。
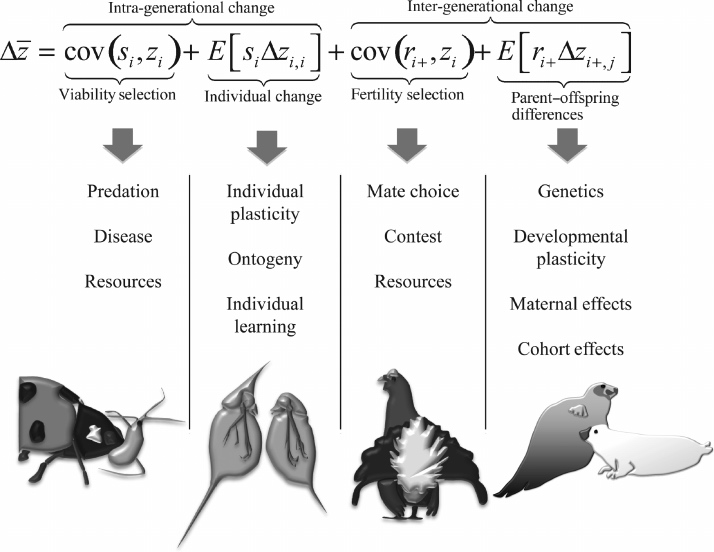
图 13:使用Price方程分解表型变化的迭代演化,显示了影响每个组成部分机制的例子[4]
相关理论研究和关系
对与其他以数学测量作为方法的相关研究进行总结,有益对作者理论有更好的理解。
-
作者的研究方法与 Maturana(Maturana, 1975)发展的自创生(autopoiesis)概念有关,他强调在自主性方面,从事自我生产的过程网络的“统一”(Maturana, 1980),以及格式塔感知的概念,在这些观念中,人物观察到的超过其各部分的综合,并与这些部分有不同的实质基础。
-
另一个相关的工作是对模块化网络科学的研究。对于静态结构,有合理的模块化定义,大多都与将微观数据划分为更具紧密约束的群集(如群落)过程有关。例如,在网络中,量化模块度量往往寻求对节点和边的集合划分,与适当的空模型相比,这些集合在数据统计中是被过表示的(overrepresented)(Newman, 2016)。至于发展性的模块定义,例如那些应用于胚胎发育、器官形成的理论,也为理解个体性提供了一个视角 (Davidson et al., 2004) ,只是它们还没有像网络科学中那样以定量测量的形式识别呈现。
-
本文的研究与 Karl Friston 和同事 Ramstead 等人(2018)提出的自由能量原理(FEP)也有联系。与个体信息理论一样,FEP 也继承了薛定谔思想从第一原理出发,以解释自适应系统如何跨越时间抵抗衰变以实现持久性。它也强调减少不确定性,只不过是通过最小化自由能的视角来进行。自由能量原理的基本思想是,自适应系统将总在可能相空间中占据一个较小的有界状态集。此外,自适应系统通过建构将生物体与环境分开的区隔来实现自由能最小化——在自由能量原理的表述中,这个“过滤器”是一个马尔可夫毯(Markov blanket),它规定了内部和外部状态的条件独立性:内部状态只通过马尔可夫过滤来感知外部状态。
个体信息理论(ITI,the information theory of individuality)是基于第一原理的数学形式理论,通过捕捉从过去到未来的信息流,从而能够严格定义不同形式的个体。基于此,许多现有的生物概念(例如,协调的的复制体、发育中的个体),都将通过这个过程被识别为可感知的个体。如前所述,不仅包括更大时空尺度,许多在社会层面被贬低为低级形式衍生或表象的新奇 “个体 “也可能会被识别出来。此外,对于非生物现象,如而那些相对于自身动态历史、经历了相对较长的环境历史迅速涌现的预适应组织(如自组织结构,如被环境决定测量所决定的漩涡),这项研究也有很大借鉴意义。
但研究目前只是刚刚开始,一方面在选择尺度和层次上需要进一步探讨,另一方面缺少系统间的成本函数建立一个终止的阈值以获得确切边界,还没有找到一个最优的个体归纳算法, David Krakauer 表示这个问题将留给以后的工作来解决。
对几种不同类型个体和性质的细致探讨,有助于我们理解人类和自然之间的关系,尤其是人类社会中个体的独特意义。
回到前面对个体与环境之间三个耦合参数讨论:𝛼𝑆 代表个体自身系统状态之间耦合参数,如生物各种内部调节系统。𝛽𝑆 代表个体与环境交互作用的耦合参数,如生物的自适应性能力。至于𝛾𝑆,则是调节原个体系统与环境状态综合影响的参数。是一种高阶相关性。如并非直接和生物交互作用的自然选择。
在原文中,对于 𝛾𝑆,只暗含了一种情况,即个体系统与环境状态之间的高级相关性,是环境调节个体系统,最典型是自然选择,仿佛一只看不见、神秘莫测的手,主宰所有生物的演化。
然而对于人类而言,不管地球是不是已经进入所谓人类影响地球地质演化的人类世,有一点是可以肯定的,那就是地球上已经存在了另一种高阶相关性,即人类文化对自然的认知和影响。这种人类对环境反向作用,自从智人开始诞生就已经存在了。
可能是由于新冠病毒的影响,今年初在 Nature Reviews Earth & Environment [5]发表了几篇有关地球系统科学文章,讲了整个学科历史和建立。其中有一篇给出了最新版本的地球系统内外部结构作用图。
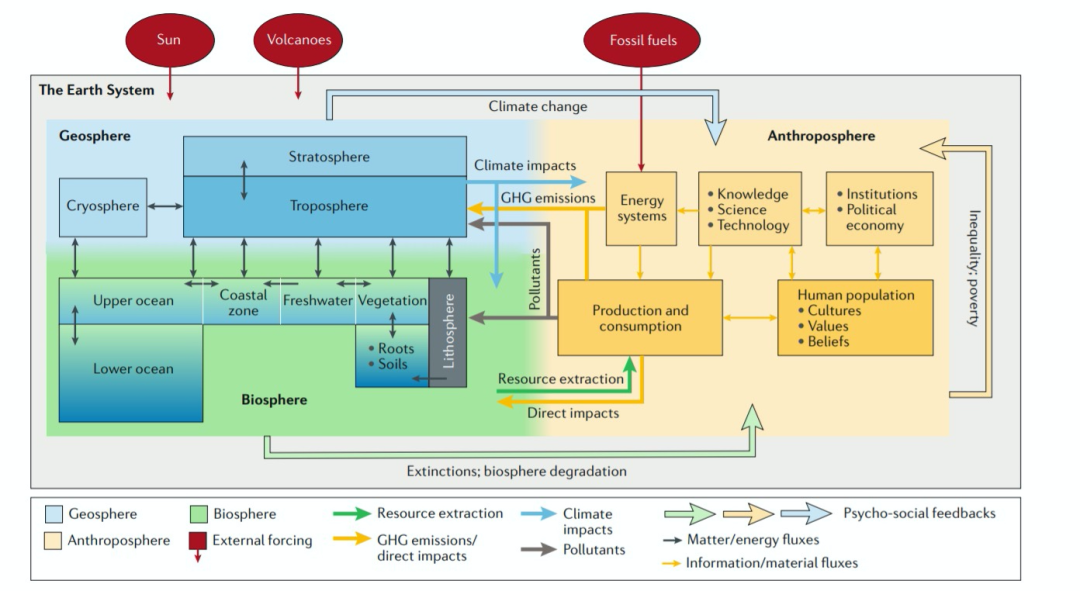
图 14:地球系统作用图
从图中我们可以看到,以三种颜色分别代表的地圈、生物圈、人类圈处于相互作用中。人类的基础设施建设、能源利用等活动都直接与地球系统相互作用并施加影响,在间接上则来源于人类的文化,包括科学与技术、人类价值以及政治、经济活动等。
结合前面的分析,一般的有机生物体,仅仅维持高 𝛼𝑆,即试图以自身状态主导自身发展时,必然会趋于灭亡,为此唯有提高与自然的适应性的 𝛽𝑆 以及 𝛾𝑆,而这会势必导致群落个体的出现,即社会属性增加。
然而,由于人类建造了城市,从而能让自身维持高 𝛼𝑆 情况下与自然环境不再直接相关,在新环境下让个体一定程度避免上了大自然的弱肉强食(当然种内社会竞争是另一回事),能够继续自身主导自身。甚至在某种意义上说,网络社会和虚拟现实依然符合这个趋势。这就意味着,人类的演化是尽量保留有机个体特性的方向的。
但同时人类和自然环境又是具有高阶相关性,人类文化对自然的认知和影响,出现了另一种情况下的 𝛾𝑆 增加的高耦合,从而导致了人类的群落化。
这种群落个体的特性,即所谓的全球化进程,让人类能在面对更广阔自然环境时,如宇宙尺度的空间和时间时,具有更强的适应性。当 𝛾𝑆 增加时,生命体为了生存,无论是个体系统还是环境哪方为主高耦合都会出现群落个体,然而这两种群落个体实质上是不一样的:如果自然环境下自然选择占据主导,就会出现如蜜蜂和蚂蚁那样的真社会性,因适应环境变得高耦合;而但如果物种自身占据主导,例如人类的知识和文化,能通过科学和技术探索和改造环境,甚至诗歌艺术吟唱大自然的秘密,就会是另一种高耦合下的——是人类以自己的精神给物质自然赋予意义的意义下的高耦合。
科学发现自然规则,诗歌为人类立法——在这种情况下,不仅需要保留有机个体特性,珍视个体价值,甚至可以说具有独立自主性的个体,是人类这种新型群落个体的条件也是充分条件。
由此可见,人类是保留了有机个体自主性的新型群落个体。用社会生物学来说,即既非前社会性(独居,Presociality),也非真社会性(Eusociality),在人类组织形式历史中,往往是分布波动在亚社会性到准社会性之间,在全球化之前,国家的产生和壮大只是进程一部分。
亚里士多德说「喜爱孤独者,非神即兽」,真社会性在某种意义上就是一个更高层次的大神(大兽,利维坦)。但如前所述,由于人类个体极高的复杂度(人脑、意识),对自然探索和文化创造本身的需要,以主动姿态和自然达成高耦合,像蚂蚁那样完全确定分工和阶层的真社会性和人类群落个体的社会性并不相同。

图 15:社群主义 VS. 自由主义
至于自由主义和社群主义之争,现在我们也有了全新理解。从对人类个体和新型群落个体的分析可以看出,注重个体价值不是意味着要滑向唯我论和相对主义,而是让人类具有选择从非前社会性到真社会性的能力,从而具有超越两极最大的适应性:在保证个体价值性特殊的情况下,让人类具有了从每一个个体思考预演整个人类命运的能力,都参与到人类这个整体生命过程中来。
——区别于所有其它动物,人类是同时向所有可能性道路敞开探索,只要不是所有个体都是错误的,人类在宇宙延续就总有希望。每个人都是希望的一部分,关心自己,就是关心他人,关心人类本身。
这可能就是人类作为个体生命最特殊的意义。
[1] 部分信息分解(the partial information decomposition)(Williams & Beer, 2010) 这个术语,是从一组普遍公理中推到出来的。但这些公理并没有为特定术语定义其具体的度量。我们需要对这四个术语中的一个进行额外定义。Williams 和 Beer 最初的研究建议(2010)被批评为反直觉,其后有一系列其他的研究建议 (Harder et al., 2013; Bertschinger et al., 2014; Finn and Lizier 2018; James et al., 2017),不过尚未达成共识。在本文中,把“分解”作为一个概念框架,来解释经典信息论的度量。
[2] https://globalnews.ca/news/6799525/jellyfish-string-creature/
[3]https://www.researchgate.net/publication/230758418_Understanding_behavioural_responses_and_their_consequences
[4] https://www.nature.com/articles/s43017-019-0005-6
作者:十三维
审校:刘培源
编辑:张希妍
推荐阅读

集智俱乐部QQ群|877391004
商务合作及投稿转载|swarma@swarma.org
◆ ◆ ◆
搜索公众号:集智俱乐部
加入“没有围墙的研究所”

让苹果砸得更猛烈些吧!
👇点击“阅读原文”,了解更多论文信息
微信扫一扫,分享到朋友圈











