从这篇总结性文章中,我们将一起了解:连续定向进化系统的突破点是什么?连续定向进化系统分为哪些种类?各自的优劣势又有哪些?连续定向进化系统的启示是什么?
孟凡康 | 作者
再创 | Regenesis | 来源

论文标题:
Systems for in vivo hypermutation: a quest for scale and depth in directed evolution
论文地址:
https://www.sciencedirect.com/science/article/abs/pii/S1367593121000302
今天我要和大家分享的一篇文章是近期发表在Current Opinion of Chemical Biology上的文章「Systems for in vivo hypermutation: a quest for scale and depth in directed evolution」,通讯作者是美国加利福尼亚大学尔湾分校的Chang Liu。如果各位读者了解定向进化的话,Chang Liu的名字应该会经常出现,该研究组最著名的工作之一是细胞内的连续定向进化系统—OrthoRep系统(本文会有介绍)。
-
-
连续定向进化系统分为哪些种类?各自的优劣势又有哪些?
-
进化是终极的生物工程师,但复杂的基因结构正在让生物的进化步伐变慢。
从生物单个基因的角度来看,进化的步伐在40亿年后已经变得非常缓慢。这不仅仅是一个经验性的观察,而是生物系统中日益复杂所带来的基本结果:随着生命体变得越来越复杂,即使微小的变动也可能会在复杂体系中打破更多原有的结构。因此,基因组的突变率必须保持在低水平。
但对一个蛋白质工程师来说,这种结果会产生一个大麻烦:如果编码蛋白质的基因必须服从基因组的低突变速度限制,在细胞内对任何特定基因的进化将无法快速推动,复杂蛋白质功能的开发也将受到极大的限制。
近些年,连续定向进化系统成为一个迅速发展且受到持续关注的研究领域。简单来说,其目的是设计和应用遗传系统,在细胞内选择性地对目标基因(Gene of Interest,以下简称GOI)进行高效进化。这种系统可以绕过基因组的低突变速度限制,推动GOI的快速连续进化。
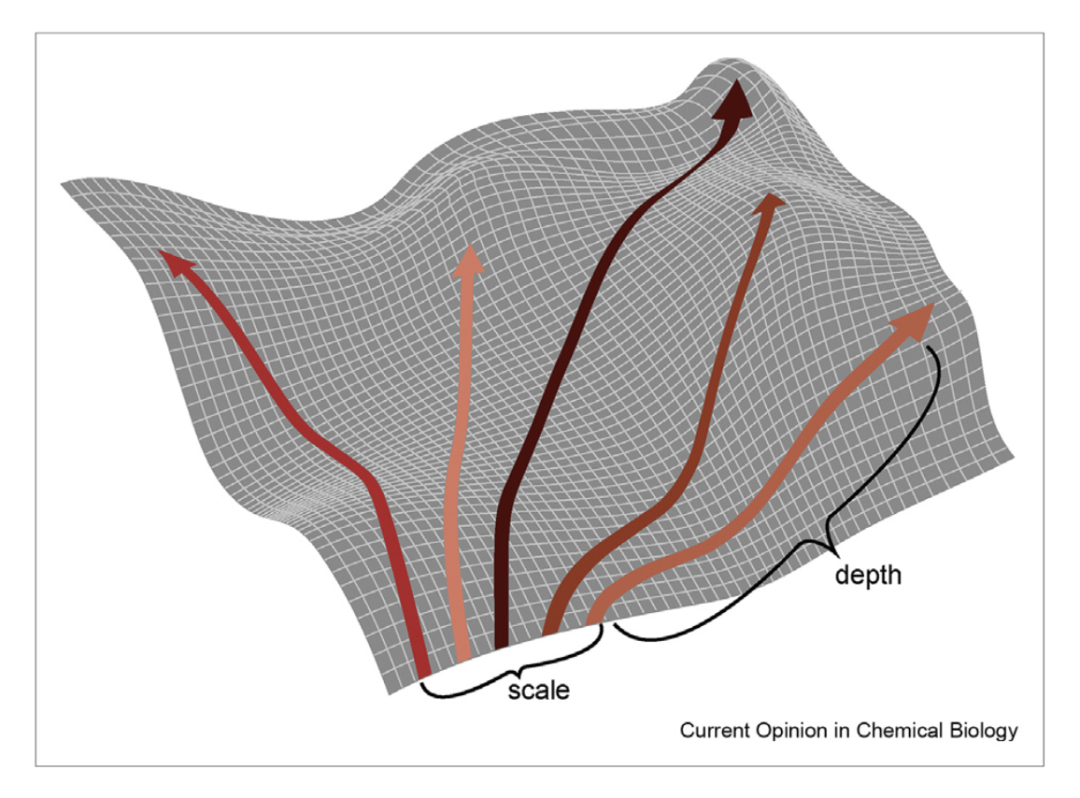
生物本身就是一个自动化系统,可以在「深度」以及「规模」上超越传统定向进化系统。
不过,成熟的体外定向进化已经使在实验室时间尺度上进化GOI成为可能,为什么还需要细胞内的连续进化系统?
在经典的定向进化中,研究人员对一个或多个GOI进行体外突变建库,然后将突变库转化到细胞中,并对所需功能进行筛选或选择。这个过程可以被看作是手动绕过基因组突变速度限制的过程。然而,经典的定向进化在很大程度上放弃了自然进化的两个关键性的优势:「规模 」和「深度」。
第一,规模(进化数量受限)。突变、转化和选择的人为操作具有劳动密集型的特点,同时在技术上也通常具有挑战性。因此,在一次尝试中很难实现进化实验的规模化。规模太少很容易陷入局部最优解的困境。
第二,深度(迭代次数不足)。因为突变、转化和选择的人工周期可能需要几天到几周,所以研究人员无法在适应性景观上进行长距离的摸索。但诸如探究复杂的选择和适应性之间的关系、实现复杂的蛋白质进化结果(新的酶活性,或复杂的蛋白质-蛋白质或蛋白质-核酸相互作用),都需要在适应性景观上进行长的突变路径摸索。
然而,同时具有「规模」和「深度」的过程在物种的自然进化中却是大量存在。所以,建立能在体内实现GOI定向突变的遗传系统的关键动机是将「经典的定向进化转变为为自主和连续的过程」,从而突破传统进化无法达到的「规模」和「深度」。
到目前为止,连续定向进化系统一般通过三种框架来实现:
-
病毒框架: 病毒基因组在非扩增性宿主中进行突变富集。
-
-
直接突变框架: 具有突变能力的正交DNA复制系统。
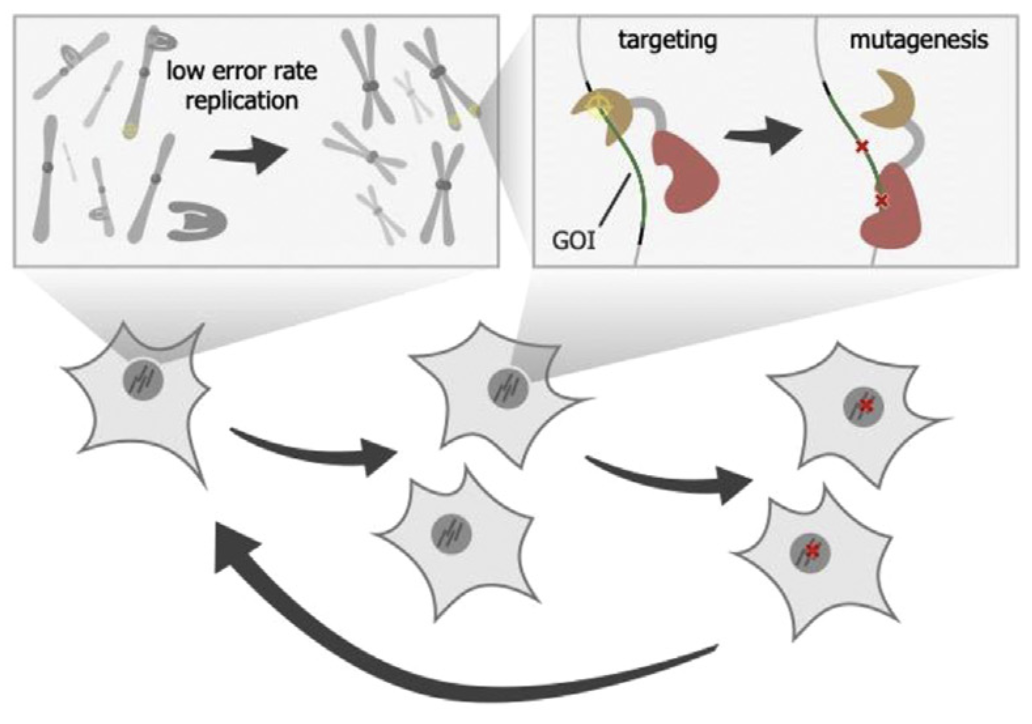
在病毒框架中,GOI被编码在病毒的基因组上,当病毒在宿主细胞中复制时会被诱导产生突变。GOI的表型同时会与病毒复制的能力建立关联:所以当病毒利用宿主不断复制迭代时,具有优势的突变体会逐渐在群体中富集。这便是噬菌体辅助持续进化(PACE)系统背后的基本策略,它是病毒框架的最成熟的例子。
在PACE中,GOI被编码在M13噬菌体基因组上,噬菌体通过大肠杆菌宿主细胞繁殖。大肠杆菌本身的DNA复制系统通过改造,能够快速诱导噬菌体变异。尽管高突变率(在每碱基10^-5到10^-4个替换)对大肠杆菌是有害的甚至是致命的,但这对该系统来说不是问题,因为大肠杆菌在PACE系统中会不断流入和流出含有噬菌体的培养池,只有噬菌体群体在PACE系统中是持续存在的。突变只会在噬菌体基因组和GOI中不断积累(因为噬菌体基因组较小,在GOI之外的突变积累造成的问题不会很大,当然这是一个隐患)。
pIII蛋白是噬菌体包装和感染的必需基因。一般来说,pIII蛋白越多,噬菌体的后代就更多。在PACE中,M13噬菌体基因组缺少gIII基因,但gIII基因存在于大肠杆菌宿主中,其gIII基因的表达与GOI的表型相关联。只有符合选择表型的GOI突变体才会诱导gIII基因的表达。组装完成的噬菌体会裂解宿主,侵染新的宿主。而不符合预期表型的GOI突变体会造成噬菌体无法正确组装,从而无法裂解宿主,最终随着宿主流出PACE的培养池。这样一来,最终培养池中存在的噬菌体都将是符合选择表型的GOI突变体。
总体来说,「病毒框架」的优势在于长时间的连续或半连续的推动GOI的进化迭代,从而实现理想的进化「深度」。同时这种进化系统在扩大「规模」上也相对容易。
当然,「病毒框架」也存在诸多限制。首先,该系统受限于病毒的生理限制。病毒并不是自主繁殖的媒介,必须依赖于新宿主细胞的持续供应。这就引入了对生物反应器的要求。在PACE中,新宿主细胞的供应速度需要精确的外部控制,以便噬菌体和宿主可以及时分离(噬菌体的分裂速度稍大于大肠杆菌的分裂速度),防止宿主本身的突变影响进化结果。
其次,表型的筛选目前只能与病毒繁殖相关联。尽管我们可以通过基因线路设计将多样的GOI功能与病毒繁殖关联起来,但超越病毒生命周期尺度的功能、涉及复杂的宿主生理学的功能、或旨在改变宿主细胞(甚至是多细胞生物体)生理学的GOI功能都不能通过PACE等「病毒框架」进行直接选择。
另一个可行的方案则是在细胞内直接进行进化。但是细胞的突变率非常低,这种低突变率(通常为每碱基10^-9到10^-10个替换)不足以在单个GOI水平上快速实现突变的多样化。为了满足进化所需要的规模和深度,我们必须设计出在细胞内有针对性的突变系统来加速GOI的进化。
一个简单的策略是将DNA突变酶与具有位点特异性的DNA结合蛋白融合。比如我们可以将DNA脱氨酶与T7 RNA聚合酶进行融合—其中T7 RNA聚合酶的作用是带着脱氨酶在GOI的DNA上滑动,脱氨酶在此过程中会诱导DNA突变。同样的原理,低保真的DNA聚合酶与Cas9的融合也可以使靶向的序列附近的DNA发生突变。
这些涉及DNA突变酶与DNA结合蛋白融合的系统可以归类为「表观框架」。大家可以发现,「表观框架」中突变过程是独立于正常的DNA复制的:GOI仍然是可以通过细胞内的固有DNA聚合过程实现复制,但是在此之外,额外的一个靶向系统会造成目标基因的突变,并通过固有的DNA聚合过程积累突变。
但是「表观框架」的限制也很明显。一个问题是当一个GOI成为突变的目标时,负责招募突变机器的DNA结合序列可能会被突变机器或者宿主本身破坏掉,造成进化的速度随着时间的推移而减缓。另一个问题是目前的「表观框架」也有明显的脱靶活性,导致宿主基因组的突变率升高,可能会造成宿主通过其他方式避开选择压力。
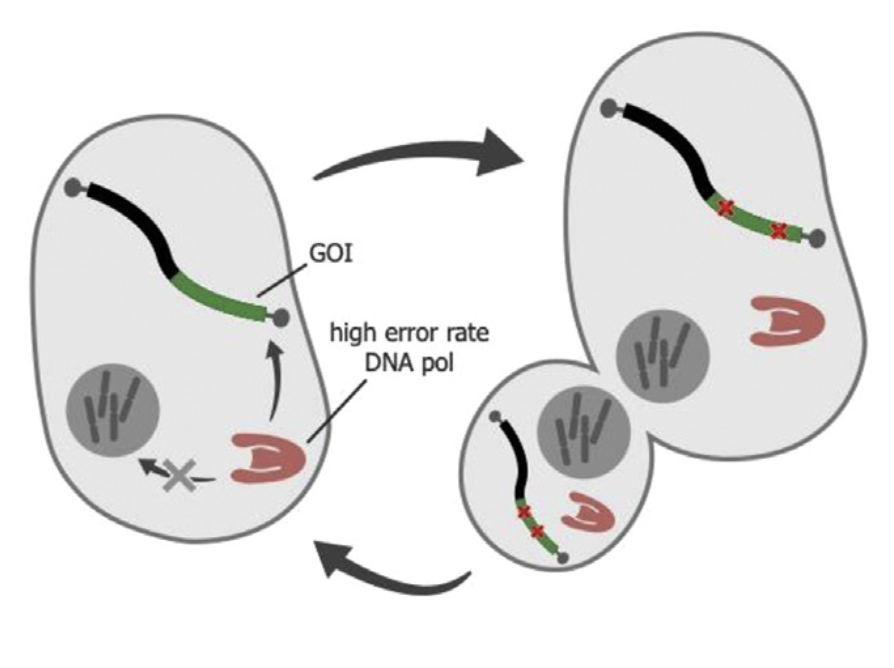
在体内实现GOI定向突变的另一个方法是「直接突变框架」—给细胞提供一个正交的DNA复制系统,专门用于GOI的复制。「直接突变框架」中的正交DNA复制系统通常包括一个特殊的低保真DNA聚合酶,它只复制和突变编码GOI的特殊载体(如质粒),推动其在细胞内快速进化。而宿主DNA聚合酶只将复制宿主基因组,不复制特殊的基因载体,实现正交性。
华盛顿大学的Lawrence Loeb等人通过建立针对ColE1质粒易出错的DNA聚合酶I,开展了建立正交复制系统的早期工作。然而,由于DNA聚合酶I对基因组DNA的复制也是必不可少的,因此没有实现完全的正交性。另一个例子著名的例子则是OrthoRep系统。研究人员发现在某些酵母菌株中存在自主复制的细胞质质粒。Liu Chang等改造了这种特殊的细胞质质粒,开发了负责复制这种质粒的DNA聚合酶的低保真版本,最终设计了OrthoRep系统。OrthoRep系统的质粒只在细胞质中复制(突变率为每碱基10^-5个替换),与细胞核中的复制过程完成正交(突变率为每碱基10^-10个替换)。
OrthoRep系统并非没有局限性。首先是其突变率相对较低,需要开发更高突变率的正交DNA聚合酶进一步加快GOI的进化速度。正交DNA聚合酶的也需要改善对DNA突变的偏好性。其次是正交质粒上的基因线路设计需要优化,提高可进化的GOI范围和特性。还需要将任意所需的GOI功能与细胞适应性结合起来,满足选择的需求。最后是OrthoRep系统的可移植性需要测试,目前OrthoRep系统只能在酵母系统中实现,能否被移植到酵母以外的生物体中,还有待观察。
细胞内连续定向进化系统核心价值是其推动进化实现「规模」和「深度」的能力,从而摆脱进化的局部最优解困境,也能让我们有能力去开发更复杂的生物大分子。「规模」和「深度」上的突破也将带来高质量的进化数据,与机器学习的结合将有可能产生更大的价值。两者的结合可能会产生一个良性的循环,最终进一步加生我们对于蛋白质序列与功能关系的理解。
笔者在整理文章的时候也有一些个人的思考,写在这里,期待与读者一起讨论,即「我们还是要回归生物本身」。此处的「回归生物本身」有两个含义:
第一个含义是更复杂的定向进化需要回到宿主本身。首先,进化是一个黑箱,给与更多的可能(比如细胞内的复杂网络)往往能创造出更复杂的结果。体外的体系复杂度难以匹敌细胞内的复杂度。简单来说,在体外进化中,现阶段往往集中在单个分子的层面,但是利用定向进化很难筛选出有预期性能的基因线路,对振荡器等动态线路的筛选则要复杂得多,更不用提复杂度更高的网络结构了。 其次是合成生物学应用场景的问题,体外或者通用底盘的进化结果不代表在宿主中或者实际环境中应用仍能展现出同样的结果,在满足应用场景的宿主中直接进行定向进化是一个必然趋势,这也是OrthoRep等系统面临的移植性的难题。
第二个含义是现阶段的生物设计仍需要从自然中寻找灵感。笔者认为,在工程生物学发展的现阶段,由于对自然生命系统的理解受限,往往难以设计出超越自然范围的生物设计。前些年与中科院某位院士一起讨论课题,我给这位院士讲解了「非天然氨基酸」系统(由Peter Schultz等推动)的设计。再介绍完设计之后,这位院士提到:为什么一定要「非天然氨基酸」这个词呢?非天然意味着自然不存在,是完全人工设计的,但是很难说其他的生物没有使用过这种所谓的「非天然氨基酸」。事实的确如此,一些古菌就在使用「非天然氨基酸」,而现有「非天然氨基酸」系统的设计其实是对古菌中直接借鉴或者后续优化。其他诸多的「非天然系统」例如人工核苷酸、镜像酶等也在最初均借鉴于天然系统。本文所提到的连续定向进化系统更是一种借鉴自然的生物设计的典型案例。现阶段,自然的精妙的生物设计足以让我们继续去挖掘和模仿,大可不必有所谓的「超越自然」以及「以前从未出现类似设计」的宣称。
REFERENCE:
1.Rix, G. & Liu, C. C. Systems for in vivo hypermutation: a quest for scale and depth in directed evolution. Curr Opin Chem Biol 64, 20–26 (2021).
集智斑图顶刊论文速递栏目上线以来,持续收录来自Nature、Science等顶刊的最新论文,追踪复杂系统、网络科学、计算社会科学等领域的前沿进展。现在正式推出订阅功能,每周通过微信服务号「集智斑图」推送论文信息。扫描下方二维码即可一键订阅: