PNAS:大脑如何整合多感官模态信息,进行因果推断?

导语
大脑作为复杂智能系统,因果推断能力是其智能的主要表现之一,但怎样在微观神经元层面解释大脑因果推断的内在机制,仍是尚未解决的难题。
例如,当我们在静止的车厢里看到旁边的列车开动,会产生自己在动的错觉。这是因为视觉系统“看到”了运动,但前庭信号却显示我们是静止的。面对不同来源的信号,大脑如何进行整合并作出合理推断?电生理记录显示,大脑神经元会编码前庭运动和视觉运动线索的组合。今年8月份发表在 PNAS上的一项最新研究通过训练神经网络模型证明,这两种类型神经元之间的权衡决定了视觉线索和前庭线索应该整合还是分离,从而帮助大脑进行因果推断。
研究领域:因果推断,多感官整合,人工神经网络

Stephanie Badde, Fangfang Hong, Michael S. Landy | 作者
十三维 | 译者
梁金 | 审校
邓一雪 | 编辑

1. 大脑如何进行因果推断?
1. 大脑如何进行因果推断?
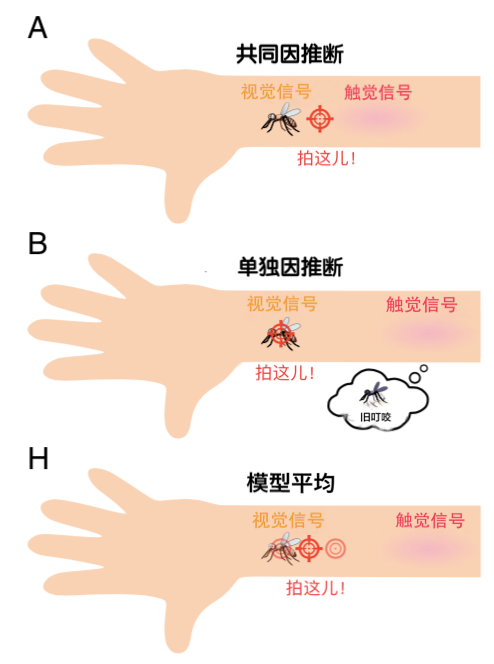
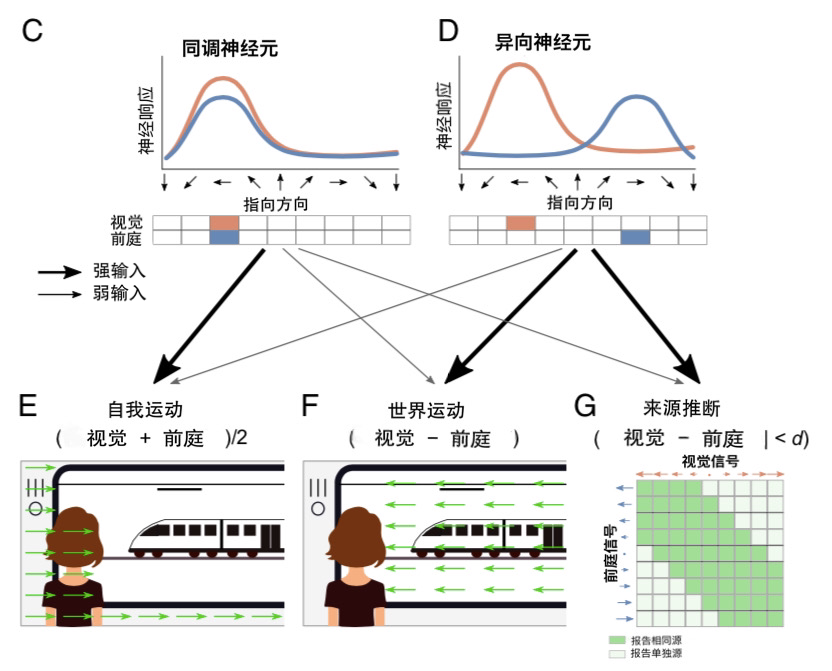
图1. 多感官整合和因果推断。(A)当推断出一个共同原因时,视觉和触觉的感觉信号被整合;(B)当推断出单独的来源时,使用分离的视觉信号。(C)在跨模态下指向方向一致的神经元具有相似的调谐;(D)异向神经元的首选方向在不同模态下有所不同。这两种类型的神经元对(E)自我运动和(F)世界运动的估计,以及(G)因果推断判断都有贡献,但程度不同。(H)在贝叶斯估计中,整合的和分离的估计被结合起来,其权重等于每种因果情况的概率。
2. 多感官信号怎么整合?
2. 多感官信号怎么整合?
*译注:大脑中某些神经细胞会对视觉、听觉和躯体感觉刺激同时起反应。一般有 50%的细胞是单通道的,但是有超过 20% 的细胞是双通道或三通道的,可以接受多感官模态信息输入。
3. 人工神经网络模拟
3. 人工神经网络模拟
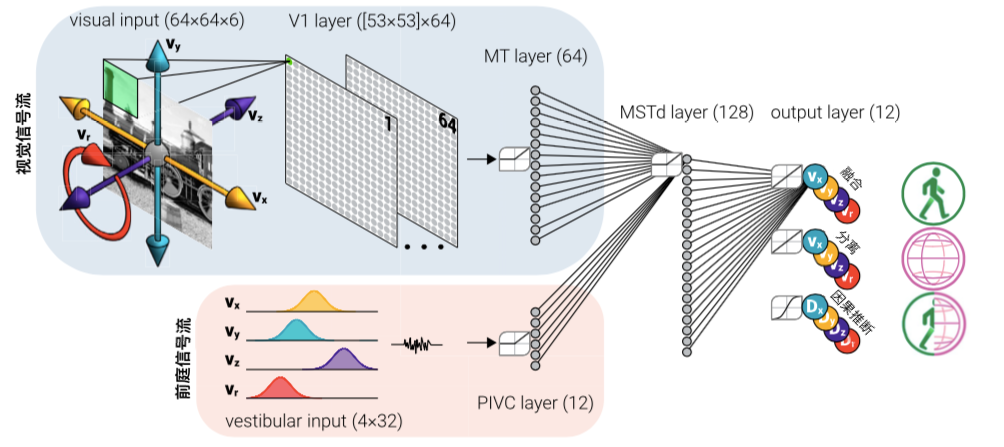
图2. 人工神经网络中的视觉和前庭信号输入,在经MSTd层三种不同方式解码后,分别输出对自我、场景和因果推断的估计
参考文献
(参考文献可上下滑动查看)
因果科学读书会第三季启动
“因果”并不是一个新概念,而是一个已经在多个学科中使用了数十年的分析技术。通过前两季的分享,我们主要梳理了因果科学在计算机领域的前沿进展。如要融会贯通,我们需要回顾数十年来在社会学、经济学、医学、生物学等多个领域中,都是使用了什么样的因果模型、以什么样的范式、解决了什么样的问题。我们还要尝试进行对比和创新,看能否以现在的眼光,用其他的模型,为这些研究提供新的解决思路。
由智源社区、集智俱乐部联合举办的因果科学与Causal AI读书会第三季,将主要面向两类人群:如果你从事计算机相关方向研究,希望为不同领域引入新的计算方法,通过大数据、新算法得到新成果,可以通过读书会各个领域的核心因果问题介绍和论文推荐快速入手;如果你从事其他理工科或人文社科领域研究,也可以通过所属领域的因果研究综述介绍和研讨已有工作的示例代码,在自己的研究中快速开始尝试部署结合因果的算法。
第三季因果科学与Causal AI读书会从2021年10月24日开始,每周日上午 10:00-12:00举办。共11-12期,每周一期。持续时间预计 2-3 个月。

报名:(长期有效)

扫码报名
第一步:扫码填写报名信息。
第二步:信息填写之后,进入付款流程,提交保证金299元。(符合退费条件后可退费。)
第三步:添加负责人微信,拉入对应的读书会讨论群。
详情请见:
因果+X:解决多学科领域的因果问题 | 因果科学读书会第三季启动
推荐阅读
点击“阅读原文”,报名读书会
微信扫一扫,分享到朋友圈











