PNAS前沿:自催化网络如何促进细菌生长?
导语
转录-翻译机器是细胞自复制的核心。细菌生长遵循简单的数学规律,这是否可以通过转录、翻译这些底层生化过程来解释?2021年7月发表于 PNAS 的论文“一个统一的基于自催化网络的细菌生长规律框架”构建了细菌的自催化网络框架,并推导说明,核糖体循环、RNA聚合酶循环等过程之间的耦合促进了细菌的均衡指数级增长。不过,通过简单的数学关系来描述细菌生理过程是“物理的”;而这背后各个生理机制之间的复杂关系则是“生物的”,理解生化过程与生长速率之间的因果关系并不容易,需要对细菌细胞内的自催化网络有更全局的理解。
研究领域:细菌生长规律、自催化循环、自复制、转录-翻译机器

刘桂 | 作者
梁金 | 审校
邓一雪 | 编辑

论文题目:
A unifying autocatalytic network-based framework for bacterial growth laws
论文链接:
https://www.pnas.org/content/118/33/e2107829118
研究发现,细菌生长的情况可以用简单的数学关系来描述,即细菌生长规律。这说明,细菌生长的核心可能存在着简单的基本原理。另一方面,细菌的细胞包含各式各样的自催化网络,以核糖体循环为例:核糖体能够翻译出组成核糖体的蛋白,而后者通过自组装形成新的核糖体。总体来说,这一过程中核糖体并没有被消耗,反而成为了自催化循环的核心催化剂。
在这篇文章中,作者构建了细菌生长的统一图景——细菌通用的自催化网络框架,并通过推导说明,自催化网络之间的关系如何促进单一细胞的均衡指数级增长。这其中的核心是转录-翻译机器,该机器包含了几个耦合的自催化循环:核糖体循环、RNA聚合酶循环、转运RNA装载循环(tRNA charging cycle)。基于该循环,作者推导循环对应的生长规律,并结合实验数据验证该规律,推测细菌的生长速率如何受温度、靶向药物等因素影响。
一、引入
一、引入
1、细胞内的自催化循环
通用构造机是冯·诺依曼设计的一种机器,它能够通过获取指令、消耗材料建造包括自己在内的一些机器,因此是自复制的。细胞作为能够自复制的个体,它的转录-翻译机器可以作为冯·诺依曼通用构造机的一个体现,而这种机器也正是自复制单细胞生物的核心。
转录-翻译机器由RNA聚合酶以及核糖体这两个关键分子机器构成。根据中心法则,细胞中的所有蛋白质均由这套核心机器分两步合成:先是RNA聚合酶转录基因、形成“指令集”mRNA;然后核糖体通过翻译mRNA生成蛋白质。
要成为一个“通用构造机”,转录-翻译机器必须能够自我复制。转录-翻译机器的自复制是个复杂、但所有单细胞生物均俱备的过程。该自复制过程由两个重要的自催化循环所构成:RNA聚合酶自催化循环以及核糖体自催化循环。如果RNA聚合酶没有转录出核糖体RNA(rRNA),细胞就不能从头合成新的核糖体;同理,如果没有核糖体翻译出RNA聚合酶蛋白亚基对应的mRNA,RNA聚合酶的从头合成也就不能实现。因此,两个循环是耦合的,且都包括自组装的步骤。
RNA聚合酶和核糖体的自催化循环也都依赖于其他的自催化循环,这些循环也是转录-翻译机器的组成部分,它们负责tRNA与氨基酸的结合、协助核糖体翻译过程的启动、移位和终止等等。所有的自催化循环相互交织,彼此需要,移除其中任何一个循环的任意一个关键的催化剂都会破坏所有的自催化循环。
2、细菌生长规律
细菌生长规律(growth law)旨在研究在忽略细胞内因相互干扰的情况下,细菌生长所表现出的整体特征与某一组分之间简单的数学关系。例如:图1a表示了一组细菌生长速率λ与核糖体质量分数(纵轴,由于核糖体RNA占RNA约为85%,因此RNA/总蛋白比率与核糖体质量分数线性相关)之间的关系:当翻译工作正常时,生长速率与核糖体质量分数呈线性正相关(用实线表示);当翻译受到抑制时,生长速率与核糖体的关系呈负相关(用虚线表示);当完全抑制时,质量分数收敛于交点处,该点表示了翻译完全受限的情况下核糖体工作的分配情况。与图1a相似,图1b表示了不同情况下生长速率与非核糖体蛋白数量(图1c中粉色的部分)的线性关系。
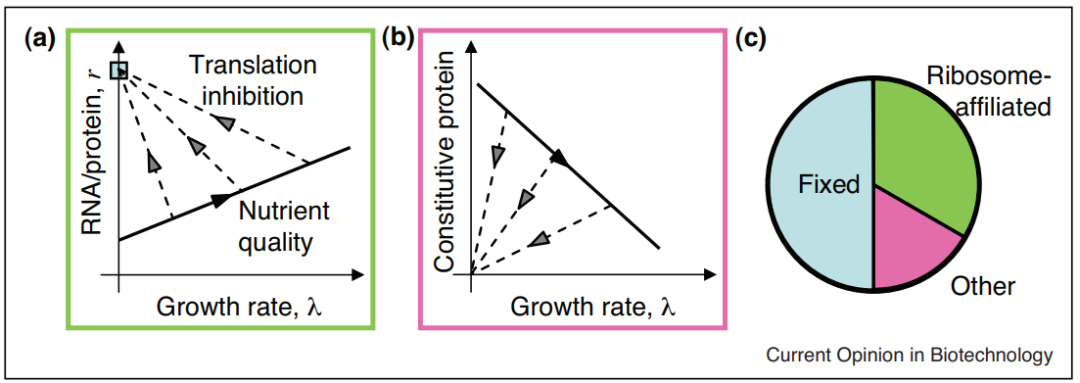
图1. 一些细菌的生长规律[1]
核糖体中心论(ribo-centric)聚焦于“核糖体制造核糖体”的核糖体自催化循环,它的生长规律研究将细菌生长速率与核糖体蛋白质质量分数、核糖体翻译速率定量联系了起来。核糖体中心论也有其短板——忽视了中心法则中的转录部分,同时也忽视了细胞内其他的自催化循环。比如,当温度或转录发生微扰时,生长速率会明显改变;而以往的核糖体生长规律却认为,该扰动并不会使核糖体的质量分数发生改变。因此,需要更合适的模型去解释这一偏差。
在这个研究中,作者兼顾了转录与翻译,推导出基于转录-翻译机器的自催化的生长规律。此外,作者发现在所有细菌内,转录与翻译过程均耦合了其他的自催化循环,而不需要其他复杂的反馈。这种耦合锁定了所有循环,使所有组分保持相同的生长速率,细胞得以呈现均衡的指数增长状态(balanced exponential growth)。
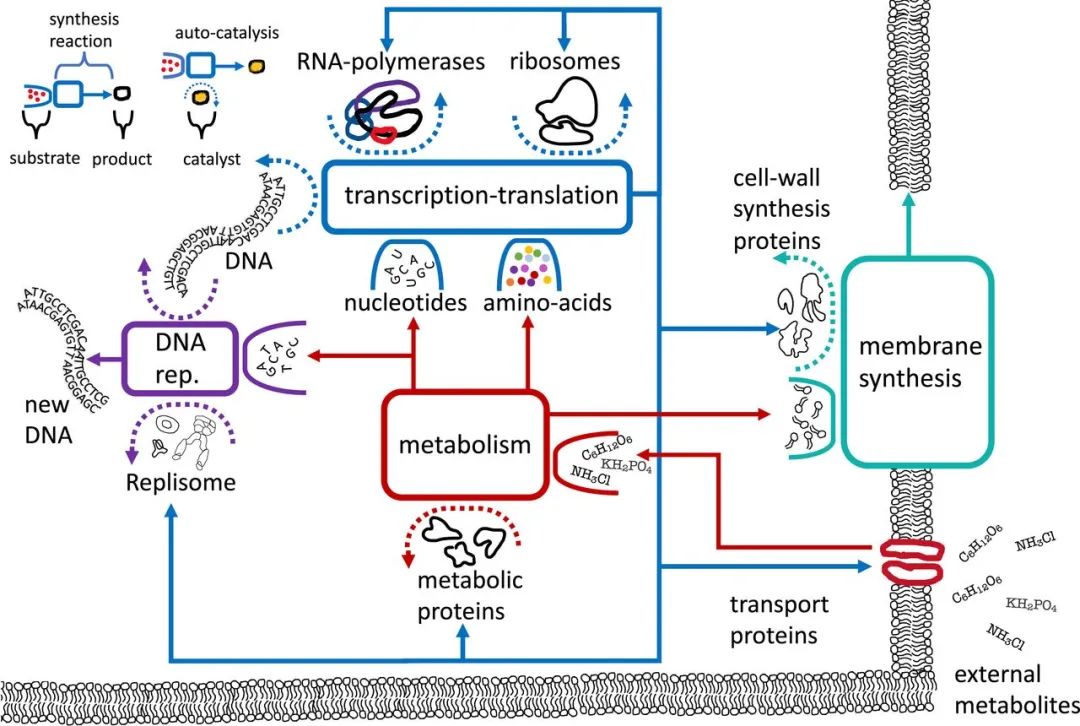
图2. 细菌的自催化网络示意图
-
方框:自催化循环的反应描述 -
实开放框:底物,被反应所消耗 -
虚开放框(含箭头):催化剂,催化反应但不会消耗 -
箭头:反应产物,流向其他的循环或退出循环
图中的自催化网络一共展示了4组耦合的自催化循环,分别是:
-
转录-翻译循环:消耗原材料和能量,产出包括自身的所有蛋白质及副本,提供大部分循环的催化剂 -
代谢循环:将外源代谢物转化为核苷酸、氨基酸、脂肪酸等代谢产物,提供大部分循环的底物 -
DNA复制循环:利用DNA作为模板复制其自身 -
膜合成循环:通过膜合成蛋白催化合成膜结构
二、结果:转录-翻译机器
利用多个耦合的自催化循环实现自复制
二、结果:转录-翻译机器
利用多个耦合的自催化循环实现自复制
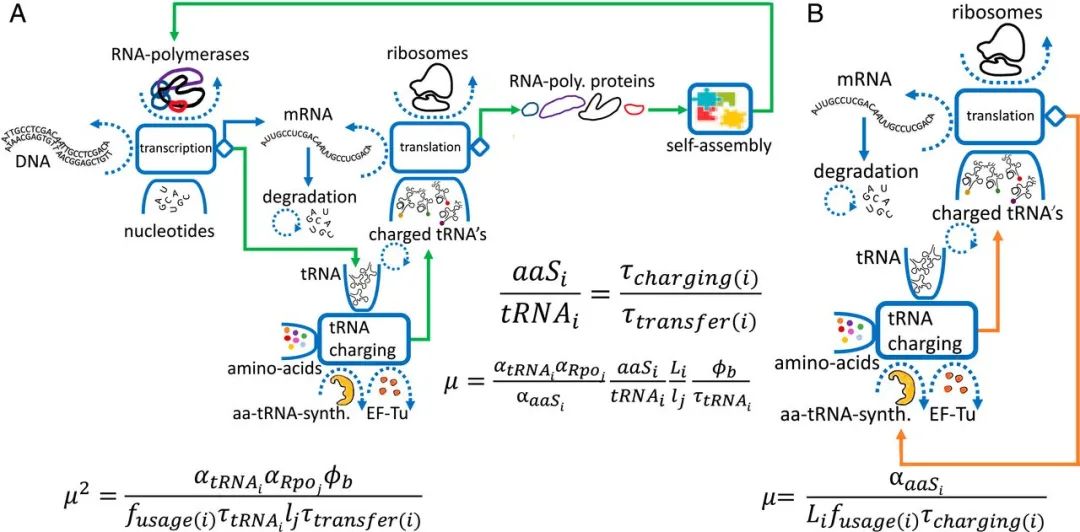
如图3所示,转录-翻译机器的自复制过程主要由三个耦合的自催化循环组成:核糖体循环、RNA聚合酶循环、tRNA装载循环,具体如下:
-
核糖体自催化循环:包含核糖体合成核糖体蛋白,RNA聚合酶合成rRNA两部分,然后核糖体蛋白与rRNA自组装形成新的核糖体 -
RNA聚合酶循环:RNA聚合酶转录mRNA,翻译产生RNA聚合酶蛋白亚基,后者经过自组装形成新的RNA聚合酶 -
tRNA装载循环:氨酰tRNA合成酶催化tRNA的氨基酸装载工作,而后tRNA运输的氨基酸参与到了核糖体的mRNA翻译过程之中,这其中就包括了氨酰tRNA合成酶的mRNA -
任何全局未被消耗的底物均可视作催化剂,如mRNA之于蛋白合成反应 -
其余与翻译相关的循环不做讨论
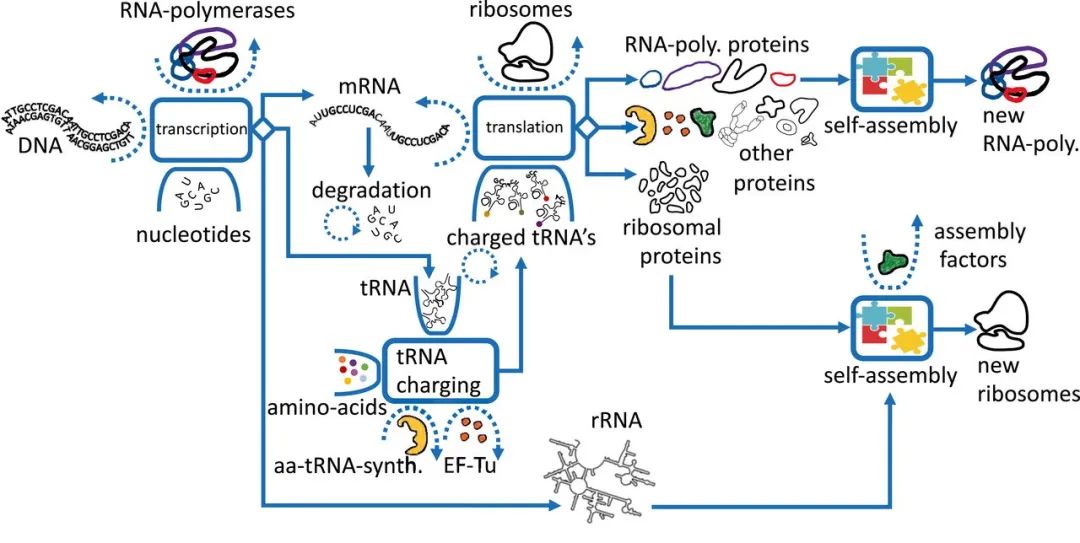
1、核糖体自催化循环


2、RNA聚合酶自催化循环

3、tRNA 自催化循环
-
tRNA自催化循环:tRNA被RNA聚合酶转录,成熟的tRNA在热不稳定延伸因子(EF-Tu)的帮助下进入核糖体;一些被tRNA运送的氨基酸最终会形成新的RNA聚合酶,其中一部分聚合酶将对应tRNA的转录,完成整个循环,tRNA生长规律见图5A下
-
氨酰tRNA合成酶(aaS)循环:在氨酰tRNA合成酶催化作用下,tRNA与氨基酸相结合,而结合后的一部分tRNA参与了新氨酰tRNA合成酶的合成,完成整个循环,氨酰tRNA合成酶生长规律见图5B下
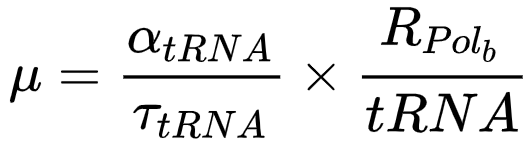
三、自催化生长规律的进一步应用
三、自催化生长规律的进一步应用
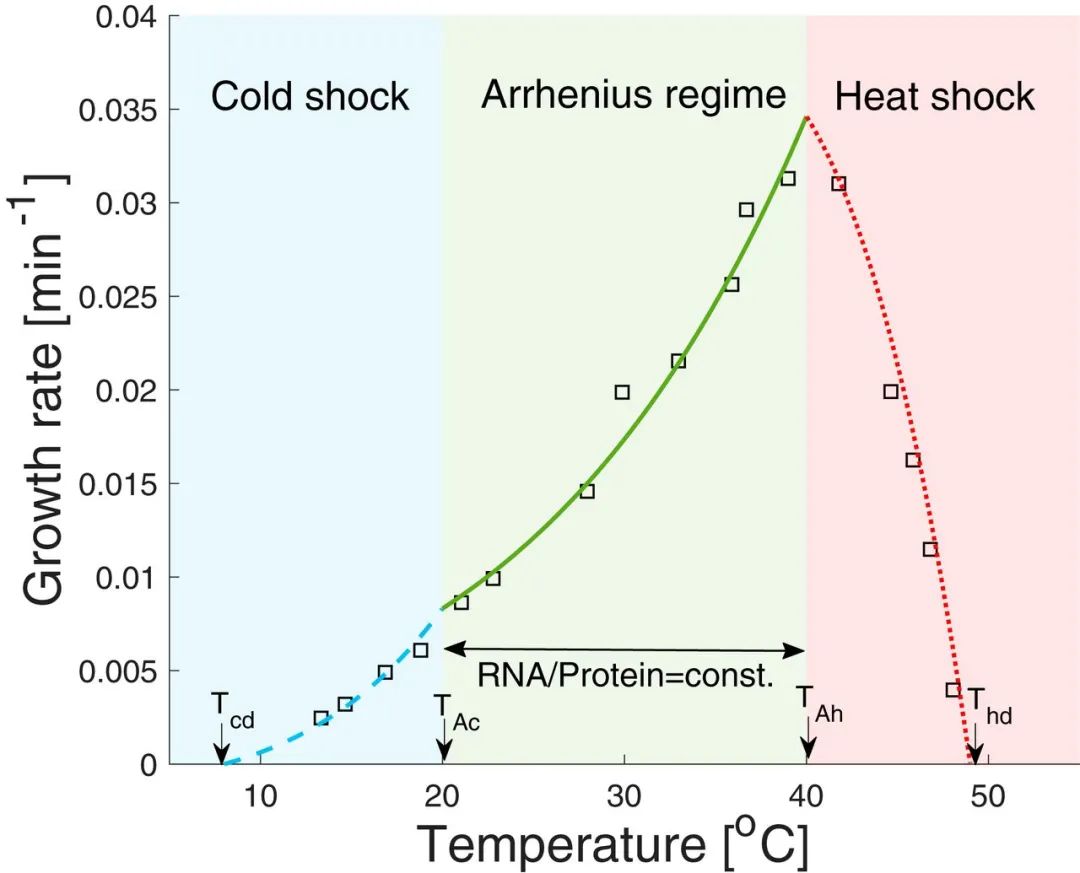
1、生长速率与温度的关系
2、抗菌药物对大肠杆菌生长的影响

 线性相关。当
线性相关。当 远小于1时,组装时间带来的影响可以忽略不计;当趋近于1时,说明因为组装时间的增加,生长速率趋近于0。
远小于1时,组装时间带来的影响可以忽略不计;当趋近于1时,说明因为组装时间的增加,生长速率趋近于0。
四、讨论延伸
四、讨论延伸
[1]. Scott M, Hwa T. Bacterial growth laws and their applications. Curr Opin Biotechnol. 2011 Aug;22(4):559-65. doi: 10.1016/j.copbio.2011.04.014. Epub 2011 May 16. PMID: 21592775; PMCID: PMC3152618.
自生成结构读书会报名中
生命是有心(意识)的,但当你一层层剥开生命的组织,却只有心脏而没有一颗“心灵”;打开大脑皮层看到一个个神经元,却看不到“意识”。然而,生命与意识都具有”自我生成”的能力,生命自发从非生命中生成,意识自发从生命中生成。更惊人的是,生命与意识的自我生成结构似乎很相似。如果这个假设成立,那么最可能的备选结构会是什么呢?
为了更深入地认识复杂活系统的自生成结构,集智科学家小木球(仇玮祎)联合周理乾、王东、董达、刘宇、苑明理、傅渥成、章彦博等科学哲学、计算机科学、物理学和生物化学等学科的一线研究者共同发起组织《自生成结构系列读书会》。其中第一季《共识——自生成结构与自复制自动机的研究背景》,将自2021年11月20日开始,其后拟于每周五晚19:00-21:00线上举行,预计持续12周。欢迎从事相关研究、对相关领域有浓厚兴趣的探索者报名参与交流。

推荐阅读
点击“阅读原文”,追踪复杂科学顶刊论文
微信扫一扫,分享到朋友圈











