我们通常以为,随着自然演化,人类的大脑体积不断增大。但研究显示,在新石器时代,随着社会组织变得更复杂,世界各地的人类大脑体积实际上都显著减少。大脑体积为什么会减小?更小的大脑必然更“愚蠢”吗?最近认知科学和系统科学专业的学者在 Frontiers in Neurorobotics 上发表研究,建立了一个最小化模型,用不同神经元数量的神经网络表示大脑体积,用神经熵和自由度两个指标量化表示神经活动的复杂性。研究发现,社会交互情境下具有较小体积大脑的智能体与独立情境下具有较大体积大脑的智能体的神经复杂性相当,表明体积较小的大脑能够通过社交互动来增强其神经复杂性,从而抵消大脑尺寸的缩小。
集智俱乐部联合天桥脑科学研究院等发起【神经动力学模型读书会】,将于3月19日开始,持续10-12周。期间将围绕神经网络多尺度建模及其在脑疾病、脑认知方面的应用进行研讨,读书会详情及参与方式见文末。
研究领域:神经复杂性,香农熵,互信息,非线性动力学,遗传算法
周莉 | 作者
刘刚强、梁金 | 审校
邓一雪 | 编辑
论文题目:Shrunken Social Brains? A Minimal Model of the Role of Social Interaction in Neural Complexity
https://www.frontiersin.org/articles/10.3389/fnbot.2021.634085/full
人们普遍认为,旧石器时代之前的人类大脑体积不断增大,以此来应对社会环境由简单到复杂的过程中对人类认知要求的不断提高 [1-2]。然而,有研究显示,在新石器时代,世界各地的人类大脑体积经历了显著的减少,且超出了身体大小整体减小所预期的范围[3-9]。该如何解释人类大脑体积变小这一现象呢?
最近发表在 Frontiers in Neurorobotics 上的文章通过仿真建模方法,对此问题进行探究。仿真模型用不同神经元数量的神经网络表示大脑体积,由此形成仿真智能体。用独立的仿真智能体模拟独立的社会环境,用两个仿真智能模拟交互的社会环境,用神经熵(neural entropy)和自由度(degrees of freedom)两个指标量化表示神经活动的复杂性。研究发现,交互情境下具有较小体积大脑的智能体与独立情境下具有较大体积大脑的智能体的神经复杂性相当。这表明体积较小的大脑能够通过社会互动来增强其神经复杂性,从而抵消大脑尺寸的缩小。
有研究指出,脑容量的减少,与人口密度规模的增加和社会化程度更高的生活方式的出现时机相吻合[10]。有研究认为,是包括提高大脑效率和自我驯化(自我驯化假说认为,为了减少群体内竞争,演化压力有所增加)的演化压力的增大,导致了大脑体积的减小[11-12]。基于在其他驯养动物中观察到的这一现象[13],提出了全新世人类脑容量降低的预测[14-15]。
实际上,新石器时代以来,虽然社会环境对认知的需求有所降低,但认知本身却变得更为有效和复杂[16-17]。因此,人类大脑通过借助高度组织协作的社会环境[18],能够以更小的体积处理与更大体积时复杂性相当的活动是有道理的[19]。进一步地,认知科学的最新理论认为[20],认知的基础不仅限于大脑,还可以扩展到大脑、身体和环境。生活在一个逐渐强化的社会环境中的人类在不减弱认知能力的前提下,大脑体积会变小,我们将此称为“减少大脑体积的社会脚手架”假说。
有趣的是,在对黄蜂这种社会性昆虫的研究中,发现黄蜂在从独立个体演化到社会物种的过程中,虽然大脑区域有所减小,并未发现大脑区域的减小与社会复杂程度有特别的关系[21]。因此可以得出结论:昆虫社会的演化不伴随任何新的对应神经结构的出现[22]。虽然其原因尚不清楚,但是关于昆虫群落的计算模型表明,相较于优化个体的大脑,对群体大脑的整体优化更具有优势[23-24]。同样地,在新石器时代,人类群体虽然不能再依赖遗传相关性获得群体内的支持,但能够通过扩展群体象征身份的规模(从大家族到大城镇)实现群体认同。这使得具有共同象征身份的不熟悉的人,在共同的社会道德期望下,能够相互依赖以获得支持[24]。由此,我们可以得出一个更具一般性的假说:大脑体积减小的关键因素不是社会自身的复杂性,而是减少群体内冲突,增加相互依赖性这一更普遍的原则。
然而,在大脑体积减少的过程中,是因为上述的相互依赖性导致独立个体大脑处理的活动复杂性变低,还是因为社会环境的演化为更复杂的大脑活动提供了支撑的脚手架,使得更小体积的大脑至少可以处理与更大体积大脑复杂性相当的活动呢?这两种可能性很难通过经验进行抉择,一项支持后者的研究表明,在人类演化过程中,代表神经活动水平的大脑血流速度比大脑体积增加得更快[25]。但是,仍然需要更多的理论工作来帮助我们理解大脑体积的减小。
2. 量化神经复杂性,
比较大脑体积与社会互动的影响
本文基于一个最小化认知模型[26],通过思想实验仿真的方法[27],以最简单的形式研究大脑体积与社会性在个体神经复杂性中产生的潜在作用。用不同神经元数量的神经网络表示大脑体积,用单个仿真智能体代表独立环境,用两个交互智能体表示社交环境,用神经熵和自由度两个指标分别从静态和动态的角度量化神经活动的复杂性[26,28-30]。具体地,从静态的角度,根据神经输出值计算的香农熵捕获神经活动复杂性[31-34]。从动力学角度[35],使用标准的非线性时间序列分析方法[36-38],利用互信息(Mutual information)估计合适的延时嵌入重构参数[39],从而使用最邻近方法[40]确定神经活动的嵌入维度,即神经活动的自由度。
已经有研究证明[26,38,41-43],在没有小组竞争的情况下,相较于增加神经网络内在的复杂性,智能体之间的密切交互更容易提高神经活动的复杂性。基于现有研究成果,作者提出假设:具有较小大脑体积的交互智能体,与具有较大大脑体积的独立智能体具有相同甚至更高的神经复杂性。
每个智能体的神经结构包括传感器、神经元和执行器三个全连接层。传感器层由两个具有S型激活函数的传感器节点组成;内部神经元被建模为连续时间循环神经网络(CTRNN),每个神经元的活动由标准 CTRNN 状态方程控制;执行器层由三个执行器节点组成,两个对应于用于代理运动的左右电机,一个对应于用于调制发射信号强度的声学信号发射器。所有执行器节点都是没有内部状态的 sigmoid 单元。
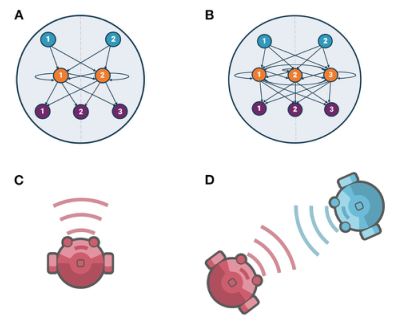
如图1所示,选择2-神经元模型(2-neuron model,有两个神经元)对应的二维动力学系统(A)和3-神经元模型(3-neuron model,有三个神经元)对应的三维动力学系统(B)进行比较研究。使用实值遗传算法优化智能体分别在单独(C)和交互情境下(D)神经控制器的参数(即连接权重、时间常数、偏差和增益)以最大化它们的神经熵。
研究比较了2-神经元模型和3-神经元模型在独立和社交两种场景的神经活动复杂性。在独立情境中的智能体称作个体演化(Individual Evolution,IE),在交互情境中的智能体称作社会演化(Social Evolution,SE)。对于IE/SE,分别对初始化的96个/对智能体进行10次独立实验,每个智能体的参数被编码为单一基因型,IE中的智能体感受不到来自环境或周围智能体的任何输入,SE中的智能体可以接受环境与其他智能体(如图1中引入的蓝色智能体)的输入并一起演化,在两种情况下,种群各自演化2000次以最大化神经熵。
图2展示了独立3-神经元模型和交互2-神经元模型的智能体的最佳演化轨迹,可以看出,交互情境下的智能体表现出比独立情境下的智能体更复杂的神经活动。在独立情境下,智能体在简单的循环中移动,在交互情境下,智能体通过螺旋嵌套的成对循环移动来增强彼此的行为。
图2. 10次仿真实验中2-神经元模型和3-神经元模型的智能体行为示意图
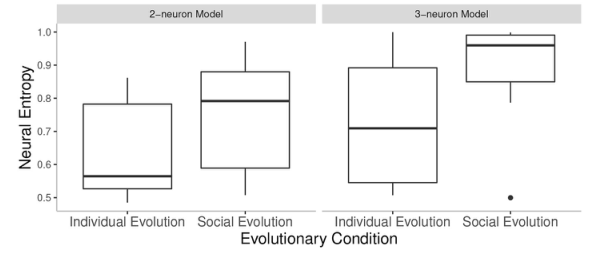
从量化的角度看, 如图3所示,无论是2-神经元模型还是3-神经元模型,交互情境下的神经熵都大于独立情境下的神经熵,3-神经元模型的神经熵明显高于相同情境下的2-神经元模型。然而,与较小大脑体积的智能体相比,较大大脑体积的智能体的神经熵并没有更大。
图3. 独立和交互环境下2-神经元模型和3-神经元模型的神经熵比较
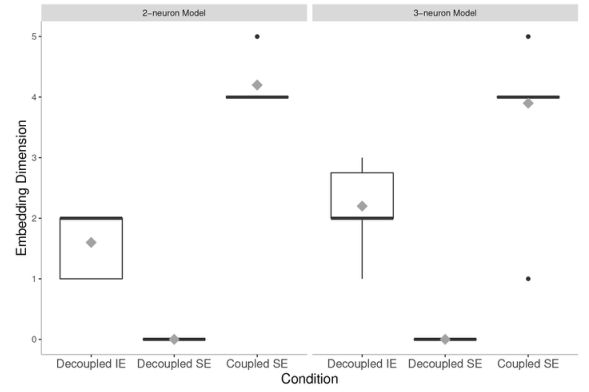
非线性时间序列动力学分析的结果验证了我们一开始的假设,如图4所示。通过比较交互情境中耦合的2-神经元模型智能体(2-neuron model, Coupled SE)和独立情境中解耦的3-神经元模型智能体(3-neuron model, Decoupled SE)的平均嵌入维度,研究发现前者具有更高的维度,即自由度更高。这证明交互的具有较小大脑体积的智能体与独立的具有较大大脑体积的智能体具有相同甚至更高的神经复杂性。
图4. 耦合和解耦情形下独立与交互2-神经元模型和3-神经元模型的神经熵
另外,从结果也可以看出,在互相耦合的交互环境下,智能体的神经活动自由度不依赖其自身的复杂性,因为在解耦时,嵌入维度会低于可实现的维度。也就是说,有可能是智能体之间神经活动的耦合增加了神经复杂性,而对于独立的个体来说,神经复杂性却在降低。
回到文章开头的问题:大脑体积的减小是因为大脑活动复杂性降低了,还是在社会脚手架的支撑下,能够以更小的体积处理与之前复杂性相当的大脑活动呢?
通过对神经熵的统计分析可以知道,大脑体积较小的交互智能体能够表现出与大脑体积较大的独立智能体相当的神经复杂性水平。这与先前的研究结论一致[26-43],说明大脑体积的减小与社会演化之间是相辅相成的,社会的高度协作与组织化为大脑体积减小提供了条件。但通过神经系统的动力学分析可以看到,在交互情境中,解耦智能体在神经状态的嵌入维数更低,这似乎佐证了大脑变得更愚蠢这一观点。
因此,在某种程度上,这两种可能性可以共存,这取决于我们关注的是大脑的拓扑结构(例如神经元数量)还是神经状态的非线性动力学(例如自由度)。因为在社会性演化过程中,个体的大脑拓扑结构可以变得简单,同时它的状态的动力学可以变得更加复杂。这就给了我们一个重要启示:在根据大脑体积推断认知能力时要更加谨慎。
1.Byrne, R.W. (1996). Machiavellian intelligence. Evol. Anthropol. 5, 172–180.
2.Dunbar, R. I. M., and Shultz, S. (2007). Evolution in the social brain. Science 317,1344–1347. doi: 10.1126/science.1145463
3.DeCasien, A. R., Williams, S. A., and Higham, J. P. (2017). Primate brainsize is predicted by diet but not sociality. Nat. Ecol. Evol. 1:0112.doi: 10.1038/s41559-017-0112
4.Wrangham, R. (2009). Catching Fire: How Cooking Made Us Human. New York. NY: Basic Books.
5.Herculano-Houzel, S. (2016). The Human Advantage: How Our Brains BecameRemarkable. Cambridge, MA: MIT Press.
6.Brown, P. (1987). Pleistocene homogeneity and Holocene size reduction:the Australian human skeletal evidence. Arch. Oceania 22, 41–67.doi: 10.1002/j.1834-4453.1987.tb00166.x
7.Henneberg, M. (1988). Decrease of human skull size in the Holocene. Hum. Biol.60, 395–405.
8.Henneberg, M., and Steyn, M. (1993). Trends in cranial capacity and cranial index in Subsaharan Africa during the Holocene. Am. J. Hum. Biol. 5, 473–479. doi: 10.1002/ajhb.1310050411.
9.Brown, P., and Maeda, T. (2004). Post-Pleistocene diachronic change in East Asian facial skeletons: the size, shape and volume of the orbits. Anthropol. Sci. 112,29–40. doi: 10.1537/ase.00072
10.Bailey, D. H., and Geary, D. C. (2009). Hominid brain evolution: testing climatic, ecological, and social competition models. Hum. Nat. 20, 67–79. doi: 10.1007/s12110-008-9054-0.
11.Hare, B., Wobber, V., and Wrangham, R. (2012). The self-domestication hypothesis: evolution of bonobo psychology is due to selection against aggression. Anim. Behav. 83, 573–585. doi: 10.1016/j.anbehav.2011.12.007
12.Hare, B. (2017). Survival of the friendliest: Homo sapiens evolved via selection for prosociality. Ann. Rev. Psychol. 68, 155–186. doi: 10.1146/annurev-psych-010416-044201
13.Cieri, R. L., Churchill, S. E., Franciscus, R. G., Tan, J., and Hare, B. (2014). Craniofacial feminization, social tolerance, and the origins of behavioral modernity. Curr. Anthropol. 55, 419–443. doi: 10.1086/677209
14.Leach, H. M. (2003). Human domestication reconsidered. Curr. Anthropol. 44, 349–368. doi: 10.1086/368119.
15.Henneberg, M. (2006). The rate of human morphological microevolution and taxonomic diversity of hominids. Stud. Histor. Anthropol. 4, 49–59. Available online at: http://www.antropologia.uw.edu.pl/SHA/sha-04-03.pdf
16.Clark, A. (2006). Language, embodiment, and the cognitive niche. Trends Cogn. Sci. 10, 370–374. doi: 10.1016/j.tics.2006.06.012
17.Sterelny, K. (2017). Artifacts, symbols, thoughts. Biol. Theory 12, 236–247. doi: 10.1007/s13752-017-0277-3
18.Sterelny, K. (2012). The Evolved Apprentice: How EvolutionMade Humans Unique. Cambridge, MA: MIT Press.
19.Bednarik, R. G. (2014). Doing with less: hominin brain atrophy. Homo 65, 433–449. doi: 10.1016/j.jchb.2014.06.001
20.Newen, A., Gallagher, S., and De Bruin, L. (eds.). (2018). “4E cognition: Historical roots, key concepts, and central issues,” in The Oxford Handbook of 4E Cognition (Oxford: Oxford University Press), 3–15.
21.O’Donnell, S., Bulova, S. J., DeLeon, S., Khodak, P., Miller, S., and Sulger, E. (2015). Distributed cognition and social brains: reductions in mushroom body investment accompanied the origins of sociality in wasps (Hymenoptera: Vespidae). Proc. R. Soc. B Biol. Sci. 282:20150791. doi: 10.1098/rspb.2015.0791
22.Farris, S. M. (2016). Insect societies and the social brain. Curr. Opin. Insect Sci. 15, 1–8. doi: 10.1016/j.cois.2016.01.010
23.Feinerman, O., and Traniello, J. F. A. (2016). Social complexity, diet, and brain evolution: modeling the effects of colony size, worker size, brain size, and foraging behavior on colony fitness in ants. Behav. Ecol. Sociobiol. 70, 1063–1074. doi: 10.1007/s00265-015-2035-5
24.Jin, K.-s., and Baillargeon, R. (2017). Infants possess an abstract expectation of ingroup support. Proc. Natl. Acad. Sci. U.S.A. 114, 8199–8204. doi: 10.1073/pnas.1706286114
25.Seymour, R. S., Bosiocic, V., and Snelling, E. P. (2016). Fossil skulls reveal that blood flow rate to the brain increased faster than brain volume during human evolution. Ro. Soc. Open Sci. 3:160305. doi: 10.1098/rsos.160305
26.Candadai, M., Setzler, M., Izquierdo, E. J., and Froese, T. (2019). Embodied dyadic interaction increases complexity of neural dynamics: A minimal agent-based simulation model. Front. Psychol. 10:540. doi: 10.3389/fpsyg.2019. 00540
27.Di Paolo, E. A., Noble, J., and Bullock, S. (2000). “Simulation models as opaque thought experiments,” in Artificial Life VII: Proceedings of the Seventh International Conference on Artificial Life, eds M. A. Bedau, J. S. McCaskill, N. H. Packard, and S. Rasmussen (Cambridge, MA: MIT Press), 497–506.
28.Cliff, D., Husbands, P., and Harvey, I. (1993). Explorations in evolutionary robotics. Adapt. Behav. 2, 73–110. doi: 10.1177/105971239300200104
29.Beer, R. D. (1997). The dynamics of adaptive behavior: a research program. Rob. Auton. Syst. 20, 257–289. doi: 10.1016/S0921-8890(96)00063-2
30.Harvey, I., Di Paolo, E. A.,Wood, R., Quinn,M., and Tuci, E. (2005). Evolutionary robotics: a new scientific tool for studying cognition. Artif. Life 11, 79–98. doi: 10.1162/1064546053278991
31.Jaynes, E. T. (1957). Information theory and statistical mechanics. Phys. Rev. 106, 620–630. doi: 10.1103/PhysRev.106.620
32.Laughlin, S. (1981). A simple coding procedure enhances a neuron’s information capacity. Z. Naturforsch. C 36, 910–912. doi: 10.1515/znc-1981-9-1040
33.Carhart-Harris, R. L. (2018). The entropic brain – revisited. Neuropharmacology 142, 167–178. doi: 10.1016/j.neuropharm.2018.03.010
34.Dotov, D., and Froese, T. (2018). Entraining chaotic dynamics: a novel movement sonification paradigmcould promote generalization. Hum.Mov. Sci. 61, 27–41.doi: 10.1016/j.humov.2018.06.016
35.Stam, C. J. (2005). Nonlinear dynamical analysis of EEG and MEG: review of an emerging field. Clin. Neurophysiol. 116, 2266–2301.doi: 10.1016/j.clinph.2005.06.011
36.Kodba, S., Perc,M., andMarhl,M. (2005). Detecting chaos from a time series. Eur. J. Phys. 26, 205–215. doi: 10.1088/0143-0807/26/1/021
37.Perc, M. (2006). Introducing nonlinear time series analysis in undergraduate courses. Fizika A 15, 91–112. Available online at: https://dk.um.si/IzpisGradiva. php?id=35975, http://fizika.hfd.hr/fizika_a/av06/a15p091.htm
38.Froese, T., Gershenson, C., and Rosenblueth, D. A. (2013). “The dynamically extended mind: a minimal modeling case study,” in 2013 IEEE Congress on Evolutionary Computation, (Cancun, Mexico: IEEE Press), 1419–1426.
39.Fraser, A. M., and Swinney, H. L. (1986). Independent coordinates for strange attractors from mutual information. Phys. Rev. A 33, 1134–1140. doi: 10.1103/PhysRevA.33.1134
40.Kennel,M. B., Brown, R., and Abarbanel, H. D. I. (1992). Determining embedding dimension for phase-space reconstruction using a geometrical construction. Phys. Rev. A 45, 3403–3411. doi: 10.1103/PhysRevA.45.3403
41.Froese, T., and Di Paolo, E. A. (2010). Modelling social interaction as perceptual crossing: an investigation into the dynamics of the interaction process. Connect. Sci. 22, 43–68. doi: 10.1080/09540090903197928
42.Campos, J. I., and Froese, T. (2017). “Referential communication as a collective property of a brain-body-environment-body-brain system: aminimal cognitive model,” in 2017 IEEE Symposium Series on Computational Intelligence (SSCI),(Honolulu, HI: IEEE Press), 863–870.
43.Reséndiz-Benhumea, G. M., Sangati, E., and Froese, T. (2020). “Levels of coupling in dyadic interaction: An analysis of neural and behavioral complexity,” in 2020 IEEE Symposium Series on Computational Intelligence (SSCI), (Canberra, ACT:IEEE Press), 2250–2256.
随着电生理学、网络建模、机器学习、统计物理、类脑计算等多种技术方法的发展,我们对大脑神经元相互作用机理与连接机制,对意识、语言、情绪、记忆、社交等功能的认识逐渐深入,大脑复杂系统的谜底正在被揭开。为了促进神经科学、系统科学、计算机科学等领域研究者的交流合作,我们发起了【神经动力学模型读书会】。
集智俱乐部读书会是面向广大科研工作者的系列论文研读活动,其目的是共同深入学习探讨某个科学议题,激发科研灵感,促进科研合作。【神经动力学模型读书会】由集智俱乐部和天桥脑科学研究院联合发起,将于3月19日开始,每周六下午14:00-16:00(或每周五晚上19:00-21:00,根据实际情况调整)进行,预计持续10-12周。期间将围绕神经网络多尺度建模及其在脑疾病、脑认知方面的应用进行研讨。
详情请见:
神经动力学模型读书会启动:整合计算神经科学的多学科方法
点击“阅读原文”,即可报名读书会