什么是生态位 | 集智百科


在生态学中,生态位是一个物种与特定环境条件的匹配。它描述了生物体或种群如何对资源和竞争对手的分布做出反应(例如,在资源丰富同时捕食者、寄生虫、病原体稀少的时候生长) ,以及它如何反过来改变这些相同的因素(例如,限制其他生物获取资源,充当捕食者的食物来源和猎物的消费者)。”构成环境生态位尺度的变量的类型和数量因物种而异,而特定环境变量对某一物种的相对重要性可能因地理和生物环境而异”。

不会飞的蜣螂占据了一个生态位:利用动物的粪便作为食物来源。
格林尼利亚生态位 Grinnellian niche是由物种生活的栖息地及其伴随的行为适应决定的。埃尔顿生态位 Eltonian niche强调物种不仅在环境中生长并对环境作出反应,而且在生长过程中还可能改变环境及其行为。哈钦森生态位 Hutchinsonian niche使用数学和统计学试图解释物种如何在一个给定的群落中共存。
生态位的概念是生态生物地理学的核心,它关注生态群落的空间格局。“物种的分布及其随时间变化的动态是由物种的特性、环境变化所决定的……以及两者之间的相互作用——特别是某些物种,尤其是我们自己,具有改变其环境和改变许多其他物种的活动范围。”居民对生态位的改造是生态位建设的主题。
大多数物种存在于一个标准的生态位中,共享行为、适应和功能特征,类似于同一广泛分类类别中其他密切相关的物种,但也有例外。非标准生态位填补物种的一个典型例子是新西兰的不会飞的几维鸟,它以蠕虫和其他陆栖生物为食,在类似哺乳动物的生态位中生活。岛屿生物地理学可以帮助解释岛屿物种和相关的未填补的生态位。
格林尼利亚生态位
生态学意义上的生态位来源于壁龛的意义,壁龛是墙壁凹槽的意思,它本身可能来源于中世纪法语单词 nicher,意思是巢穴。这个术语是博物学家罗斯维尔·希尔·约翰逊 Roswell Hill Johnson创造的,但约瑟夫·格林内尔 Joseph Grinnell可能是第一个在1917年的一个研究项目中使用这个术语的人,即在他的论文《加利福尼亚恐龙的生态位关系》中提到。
格林尼利亚生态位概念体现了这样一种观点,即一个物种的生态位是由其生活的栖息地及其伴随的行为适应决定的。换句话说,生态位是一个物种赖以生存和繁衍后代的栖息地需求和行为的总和。例如,加利福尼亚短尾叶蝉 California thrasher的行为与其生活的灌木丛栖息地一致——它在灌木丛中繁殖和进食,并通过从灌木丛到灌木丛的慢慢移动来逃避捕食者。它的“生态位”被定义为对恐龙行为和身体特征(伪装色、短翅膀、强壮的腿)与栖息地的恰当补充。

格林尼利亚生态位可以被描述为“需要”生态位,或者说是一个满足有机体生存环境要求的地区。大多数肉质植物原产于干燥、干旱的地区,如沙漠,需要大量的阳光照射。
格林尼利安生态位可以通过非交互(非生物)变量和大尺度的环境条件来定义。在这个生态位类别中感兴趣的变量包括平均温度、降水量、太阳辐射和地形方面,这些变量在空间尺度上已经变得越来越容易获得。大多数文献都集中在格林尼利安的生态位构造,往往从气候的角度,以解释分布和丰度。目前关于物种对气候变化反应的预测主要依赖于对物种分布变化的预测环境条件。然而,人们越来越认识到,气候变化也影响物种间的相互作用,从埃尔顿学说的角度来解释这些过程可能是有利的。
这种生态位视角考虑到了生态等价物和空生态位的存在。生态学上相当于有机体的是来自不同分类群的有机体,它们在相似的生境中表现出相似的适应性,例如分别在美洲和非洲的沙漠、仙人掌和大戟中发现的不同的肉质植物。作为另一个例子,大安的列斯群岛的变色蜥蜴是一个罕见的例子,趋同演化 convergent evolution、辐射适应 adaptive radiation和生态等价物的存在: 变色蜥蜴在相似的微生境中独立进化,在所有4个岛屿上形成了相同的生态群落。
埃尔顿生态位
1927年,一位名叫查尔斯·萨瑟兰·埃尔顿 Charles Sutherland Elton的生态学家给生态位下了这样的定义: “动物的‘ 生态位’意味着它在生物环境中的位置,它与食物和敌人的关系。”
Elton 根据觅食活动(“食物习惯”)对生态位进行了分类: ““Elton 专注于一个物种的龛位,因为它在食物链中的功能作用及其对环境的影响。”

德国黑森州的海狸水坝。通过开发可利用的木材资源,海狸正在影响生活在其栖息地内的其他物种的生物条件。
从概念上讲,埃尔顿生态位引入了一个物种对环境的反应和对环境的影响的概念。与其他生态位概念不同的是,它强调一个物种不仅生长在基于可用资源、捕食者和气候条件的环境中并对其作出反应,而且在生长过程中改变这些因素的可用性和行为。在一个极端的例子中,海狸需要一定的资源来生存和繁殖,但也建造水坝,改变水流在河里的河狸生活。因此,河狸影响生活在分水岭及其附近的其他物种的生物和非生物条件。在更微妙的情况下,竞争对手以不同的速度消耗资源可能导致不同物种之间资源密度不同的循环。物种不仅在资源密度方面有不同的增长,而且它们自身的种群增长会随着时间的推移影响资源密度。
埃尔顿生态位集中在生物相互作用和消费者资源动态(生物变量)的局部尺度。由于关注范围狭窄,表征埃尔顿生态位的数据集通常采用对特定个体现象进行详细实地研究的形式,因为这类生态位的动态性难以在广泛的地理尺度上衡量。然而,在解释一个物种对全球变化的忍耐力时,埃尔顿生态位可能是有用的。因为生物相互作用的调整不可避免地改变非生物因素,所以埃尔顿生态位可以用来描述一个物种对新环境的整体反应。
哈钦森生态位

这种紫喉加利蜂鸟喙的形状与花的形状互补,并与之共同进化,使它能够利用花蜜作为一种资源
哈钦森生态位是一个”N维超体积”,其中的维度是环境条件和资源,这些条件和资源决定了个人或物种实践其生活方式的要求,特别是其种群的持续生存。“超体积”定义了资源的多维空间(例如,光、养分、结构等)以及”除考虑中的物种外,所有其他物种均视为坐标系的一部分。”
生态位的概念在1957年被动物学家乔治·伊夫林·哈钦森 G. Evelyn Hutchinson普及。哈钦森探究了为什么在任何一个栖息地都有这么多种生物的问题。他的工作启发了其他许多人开发模型来解释在一个给定的群落中有多少以及如何共存相似的物种,并导致了“生态位宽度”(给定物种使用的资源或生境的多样性)、“生态位划分”(共存物种的资源分化)和“生态位重叠”(不同物种使用的资源重叠)的概念。
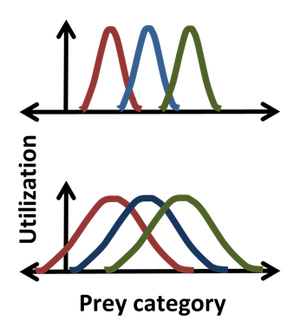
当三个物种吃掉一些同样的猎物时,每个生态位的统计图显示三个物种在资源使用上的重叠,这表明哪里的竞争最强。
统计学是由Robert MacArthur和 Richard Levins 引入到哈钦森生态位中的,他们使用直方图来描述出现的频率作为哈钦森坐标的函数。所以,比如说,高斯分布可能描述了一个物种捕食一定大小猎物的频率,给出了更详细的生态位描述,而不是简单地指定一些中位数或平均猎物大小。对于这样的钟形分布,位置,宽度和形式的生态位对应的平均值,标准差和实际分布本身。使用统计数据的一个优点在图中得到说明,很明显,在较窄的分布范围(顶部)中,极左和极右物种之间不存在对猎物的竞争,而在较宽的分布范围(底部)中,生态位重叠表明所有物种之间都存在竞争。资源利用方法假定不仅可以发生竞争,而且竞争确实发生,资源利用的重叠直接使竞争系数的估计成为可能。然而,这种假设可能是错误的,因为它忽略了每个类别的资源对生物体的影响,以及生物体对每个类别的资源的影响。例如,重叠区域中的资源可以是非限制性的,在这种情况下,尽管存在利基重叠,但对该资源不存在竞争。

作为一种半寄生植物,这棵树上的寄生槲利用它的寄主获取养分,并作为一个生长的地方。
一个不受其他物种干扰的生物体可以利用其生存和繁殖所需的全部条件(生物和非生物)和资源,这就是所谓的基本生态位。然而,由于来自其他生物的压力,以及与其他生物的相互作用(即种间竞争)物种通常被迫占据比这更窄的生态位,而且它们大多高度适应这个生态位,这被称为已实现的生态位。哈钦森把资源竞争作为驱动生态学的主要机制,但是过分强调这一点已经证明是利基概念的障碍。特别是,过分强调一个物种对资源的依赖,导致对生物体对其环境的影响重视不够,例如,殖民和入侵。
这个词是由古生物学家乔治·盖洛德·辛普森创造的,用来解释一个种群如何从一个适合它的位置跳到另一个适合它的位置,跳到一个适合它的位置,通过一些修改,或者可能是食物链的一个变化,使得适应区域对它来说是可利用的,而且它的生活方式没有中断,因为这个种群是‘预先适应’这个新的生态机会。
哈钦森的“生态位”(对一个物种所占据的生态空间的描述)与格林内尔所定义的“生态位”(一个生态角色,实际上可能被一个物种填补,也可能不被填补——见空缺的生态位)有微妙的不同。
生态位是生态空间中一个物种所占据的一个非常特殊的部分。假设没有两个物种在所有方面是相同的(称为哈丁的不平等公理)和竞争排除原则,一些资源或适应维度将提供一个特定于每个物种的生态位。然而,物种可以共享一种“生活方式”或“生态学策略”,这是生态空间的更广泛定义。例如,澳大利亚的草原物种,虽然不同于大平原的草原物种,但表现出相似的生活方式。
一旦生态位空出来,其他生物就可以填补这个空缺。例如,由于 tarpan 的灭绝而空出来的生态位已经被其他动物(特别是一种小马品种 konik)填补了。此外,当植物和动物被引入到一个新的环境中,它们有可能占据或侵入本土生物的生态位或生态位,往往比原生物种更具竞争力。人类将非本土物种引入非本土生境,往往会导致外来物种或入侵物种的生物污染。
一个物种在生态空间中的基本生态位的数学表达,以及随后生态位在地理空间中的投影,是生态位建模的范畴。
生态位和地理范围

图解表示潮间带竞争排斥对星形藤壶的影响。星斑拟南芥的基本地理分布区和实际地理分布区分别用深蓝色和浅蓝色条形图表示。
一个物种的地理分布范围可以看作是其生态位的空间反映,同时也可以看作是地理模板的特征和影响其殖民潜力的物种。一个物种的基本地理范围是它所占据的环境条件有利的区域,没有分散或殖民的障碍限制。当面对限制扩散的生物相互作用或非生物屏障时,一个物种将被限制在其实现的地理范围内,这是其更大的基本地理范围的一个更狭窄的子集。
约瑟夫·H·康奈尔 Joseph H. Connell进行的一项关于生态位的早期研究分析了限制在苏格兰库布雷岛上的藤壶活动范围的环境因素。在他的实验中,康奈尔描述了星际线虫生态位的主要特征,并解释了它们在岛屿岩石海岸潮间带的分布。康奈尔描述星际梭鱼的活动范围的上部受到藤壶在退潮期间抵抗脱水的能力的限制。种间相互作用,即与同栖藤壶物种的竞争和蜗牛的捕食,限制了分布范围的下部。通过去除竞争的巴拉诺菌,康奈尔表明星斑拟线虫能够在没有竞争排斥的情况下延长其实现生态位的下边缘。这些实验证明了生物和非生物因素如何限制有机体的分布。
决定因素
生态位的不同维度,或者说地块轴线,代表了不同的生物和非生物变量。这些因素可能包括生物的生活史、栖息地、营养位置(食物链中的位置)和地理范围的描述。根据竞争排除原则自然保护联盟的研究,没有两个物种可以长时间占据同一环境中的同一生态位。生态位的实现参数由该物种的实现生态位宽度来描述。一些被称为专家 specialists的植物和动物,需要特定的栖息地和环境才能生存,例如斑点猫头鹰,它们特别生活在原始森林中。其他的植物和动物,被称为通才 generalists,没有那么特别,可以在一系列的条件下生存,例如蒲公英。
编辑推荐
正因同根生:生态位的博弈
https://open.163.com/newview/movie/freepid=M8GAJQ1DQ&mid=M8K8RNIN3
该课程为吉林大学公开课《人文视野中的生态学》中的第六部分。
集智读书会:地球系统科学中生态学视角:适应不确定
本期分享,主讲人将以他个人的求学和科研为例,与大家分享对宏观生态学的思考与研究经验,以点带面地对当代生态学研究板块的划分做一个介绍,同时以他读博以来的研究工作为主,介绍遥感技术、时空分析、动力系统建模及多元统计分析在各类生态学研究对象中的应用。
百科项目志愿者招募



如果你有意参与更加系统精细的分工,扫描二维码填写报名表,我们期待你的加入!

集智百科平台底层维护人员招募
为了对复杂科学做一个全面且系统的梳理,提供全面、科学、客观的基础知识,诞生了集智百科。集智俱乐部组织志愿者生产复杂科学领域的知识内容,打造复杂性科学领域的百科全书。从2013年诞生至今,百科的使命依旧继续,永不完结。
为了更好的组织协调集智俱乐部社区的力量,集智百科现开放百科平台底层维护系统。
工作职责:
-
负责集智百科的业务运维工作(e.g. 权限管理,账户管理,故障排查)
-
负责集智百科的用户体验持续提升解决方案的设计和落地(e.g. 词条编辑器体验优化,系统升级)
-
负责集智百科的服务连续性解决方案设计和落地(e.g. 容灾、监控、备份与恢复)
-
负责核心系统的代码编写、系统优化以及技术架构持续改进
-
参与开发和运维规范制定,编写相关技术文档
-
能投入一定的时间和精力参与系统维护或组内讨论(>每周1小时)
-
能积极主动地深入学习系统研发与运维相关知识
-
有一定的软件研发、运维实践经验(不论技术栈)
-
Nice to have: 有以下技术实践经验:PHP, Docker, MediaWiki, 阿里云
-
投身于构建良好的学术交流氛围,改善国内科研环境,进而促进国家科技革新的伟大事业中
-
享受集智俱乐部VIP待遇(参与一年期的维护工作)
-
以练促学,践行行业最佳实践(IT行业>9年经验的前辈带)
来源:集智百科
编辑:王建萍
-
反馈:复杂系统的自我调节机制 | 集智百科 -
量子信息与量子计算预读班:追踪量子信息革命交叉前沿 -
大规模人群模拟:观察集体愚蠢与集体智慧 | 集智百科 -
行为经济学:经济系统的行为主体是否理性?| 集智百科 -
网络可控性:结构可控性与最大匹配 | 集智百科 -
《张江·复杂科学前沿27讲》完整上线! -
成为集智VIP,解锁全站课程/读书会
点击“阅读原文”,阅读词条生态位原文与参考文献
微信扫一扫,分享到朋友圈











