为了适应自然选择,大脑在信息加工过程是高效、鲁棒而经济的。然而,学界尚不清楚这些优越属性是如何在大脑中涌现的。在最近发表于 Network Neuroscience 的一项研究中,清华大学心理学系&脑与智能实验室的研究团队发现大脑的效率、鲁棒性和经济性的涌现机制可能都与渗流紧密关联。
研究领域:神经网络,渗流相变,统计物理,复杂系统


论文题目:
Percolation may explain efficiency, robustness, and economy of the brain
https://direct.mit.edu/netn/article/doi/10.1162/netn_a_00246/110014/Percolation-may-explain-efficiency-robustness-and
为了适应进化选择,我们的大脑应该有足够的效率来处理外部信息,对意外损害(例如病变)具有鲁棒性,并且在能量使用方面具有经济性。这种进化观的推论已得到众多神经科学研究的证实。
一方面,已有研究证明大脑支持神经元、环路和皮层之间的高效信息传输 [1-4]。这种信息传输效率表现为神经元通信的低时间成本或者信息的高传播能力,受到大脑连接的拓扑属性影响 [5]。另一方面,大脑被证实具有鲁棒性——一种容忍神经元或突触连接遭遇大规模破坏(例如损伤 [6-8])而依旧维持机能运转的能力 [6-13]。此外,虽然由于神经元数量巨大,大脑不可避免地需要大量能量供应并在动物体内占据很大的空间,但大脑在网络形成(将大脑网络嵌入物理空间的成本较低)和网络运行(能源利用效率高)方面被证实是经济的 [14-19]。这些经济属性被大脑网络大小、拓扑结构和突触属性所影响 [14]。
目前,学界尚不清楚大脑的这些优越特性从何而来。这些属性与大脑网络之间的密切关系自然地启发了一种涌现观的假说,即认为这些属性可能涌现自大脑连接的某些特征。近年来,这一假说的大量推论已经从不同角度得到了验证。例如,虽然最短路径有利于信息传输效率的涌现,但在网络中维持最短路径不利于经济性,所以大脑在信息传输过程中维持着“近最短路径特性”以平衡效率和经济性[2,14,20-24]。
然而,仅凭这些间接推论仍然不足以彻底证明涌现论的假说。而实验和理论研究也存在种种限制。由于在体外或体内直接观测和控制大脑效率、鲁棒性和经济性的涌现在目前的技术上不可行 [25],支持涌现假说的关键实验证据依然不明确。理论研究虽然可作为一种替代方案,但神经科学常用的模型在没有实验参考时不支持无假设的分析。囿于上述局限,大脑效率、鲁棒性和经济性的涌现假说依然受到广泛的质疑。
为了克服这些局限,生物物理期刊 Network Neuroscience 于4月15日发表了一项理论研究,证明大脑的效率、鲁棒性和经济性的涌现机制可能都与大脑网络中的渗流过程紧密关联。
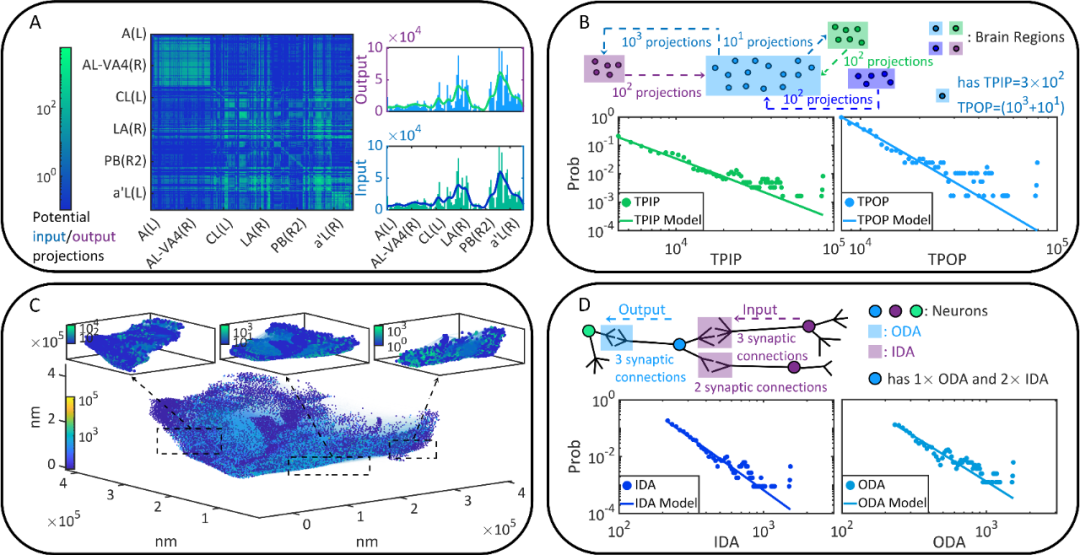
为了提升理论分析对神经科学的效度,作者使用了由FlyEM project发布的目前最大规模、最细粒度的果蝇大脑连接组 [26],能够分析微观尺度(细胞尺度)和宏观尺度(皮层尺度)的大脑静态连接(Static connectivity,即神经元网络的静态解剖结构)。分析结果见图1。
研究发现(1)微观尺度的静态连接存在更多的局部不对称性和多样的拓扑属性(不稳定的无标度性);(2)从微观跨越向宏观尺度,随着粗粒化,静态连接的对称性变强且拓扑属性逐渐趋同(稳定的无标度性)。一方面,静态连接具有强对称性,而功能连接(Functional connectivity,即神经元间信息交互形成的网络)中脑区常呈现出不对称的上下游关系,这两者之间的差异说明,功能连接的效率、鲁棒性和经济性不能用静态连接直接解释。另一方面,微观尺度的静态连接中无标度性的不稳定可能与关于大脑是否是无标度的争论 [6,27-29] 有关。
图1. 果蝇大脑的静态连接。(A) 宏观尺度的静态连接,热力图记录了不同脑区间的潜在输入、输出投射数量。(B) 对宏观尺度静态连接的部分变量的幂律分析。(C) 微观尺度的静态连接,有向图表达了不同细胞间的突触连接关系。(D) 对微观尺度静态连接的部分变量的幂律分析。
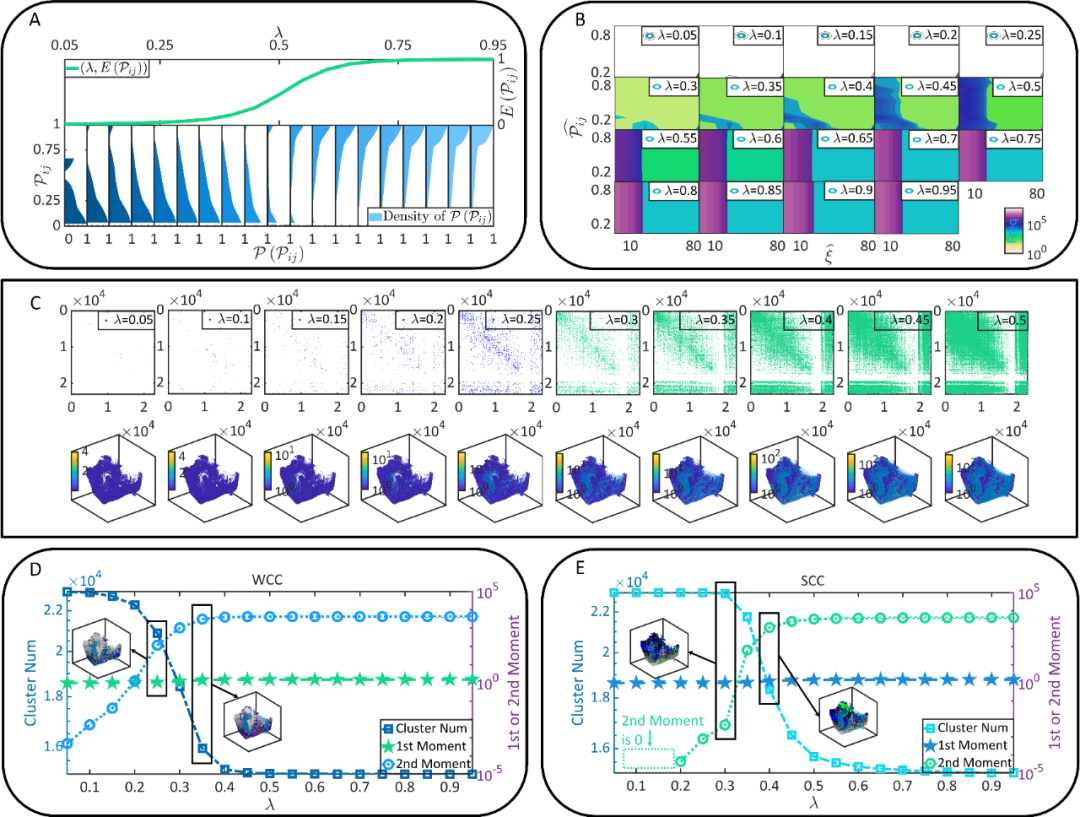
受图1结果的启发,作者发现只有理解大脑功能连接如何从静态连接上形成,才能分析大脑效率、鲁棒性和经济性的涌现。然而,现有实验技术不足以记录与果蝇大脑连接组相匹配的大规模、细粒度和高通量的神经动力学 [25,26]。为了克服这一局限,作者设定了不同的 E/I balance条件λ∈(0,1)刻画兴奋性突触连接占所有突触的比重,在不同λ条件下计算了任意两个相邻神经元的并发(coactivation)概率,从而能够分析静态连接之上功能连接的形成(功能连接形成于突触前-后的并发响应)。
在图2中,作者展示了受λ调控的功能连接的情况。上述结果从多个角度指出:随着λ的增大,功能连接开始从静态连接上形成。但当λ增大到一定程度后,功能连接的增长将衰减并逐渐被静态连接所约束。
图2. 果蝇大脑的功能连接的形成。(A-B) 不同λ条件下相邻神经元并发概率的二项分布、期望值、以及分布峰值。(C) 不同λ条件下的神经元并发情况及功能连接的形成。(D-E)不同λ条件下功能连接中弱连通分支(WCC)和强连通分支(SCC)的情况 。
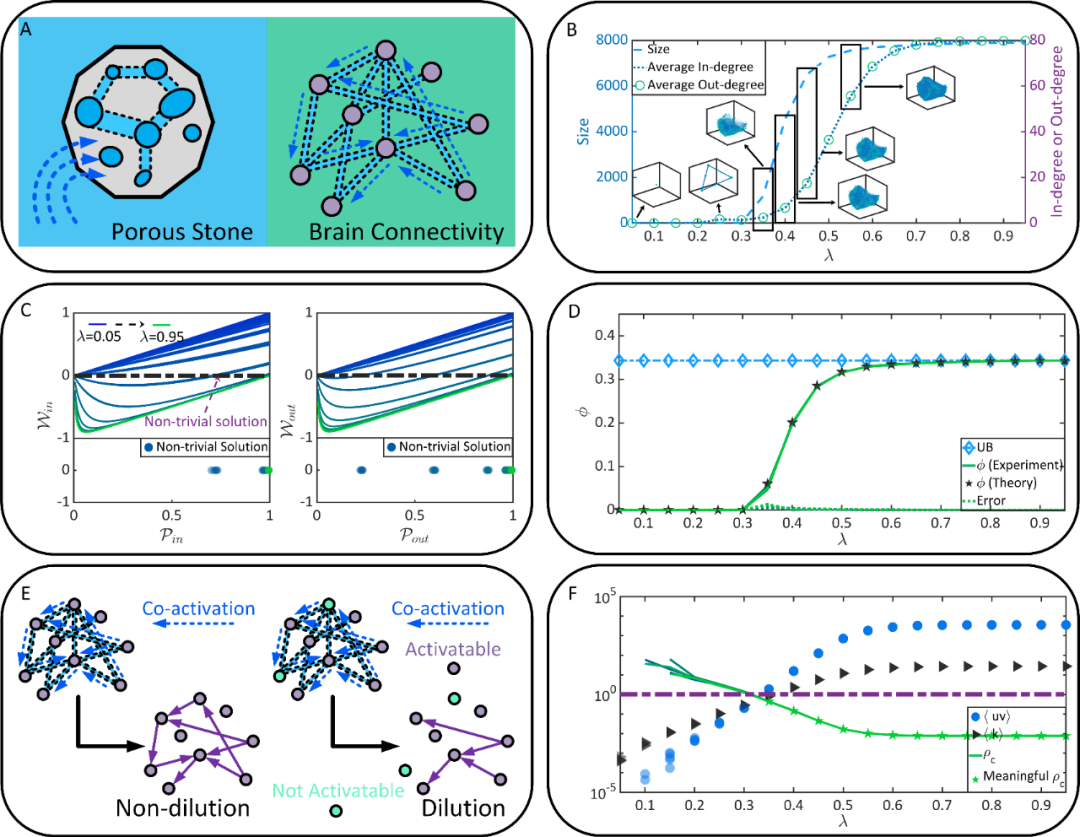
图2的结果提供了使用渗流理论分析大脑效率、鲁棒性和经济性涌现机制的机会。为了理解这一思路,读者可以将大脑想象为一块多孔的石块,神经元对应孔洞,而两个孔洞相通当且仅当两个神经元间能形成功能连接。当我们将多孔的石块浸入水中,水将流入石块并有可能连通多个孔洞——正如外界刺激可能诱发多个神经元的并发。如果暂时忽略统计物理背景,渗流(Percolation)可以简单理解为“水流是否能够贯穿整个石块”这一问题 [见图3(A)]。
显然,我们会发现如果孔洞间存在通道的概率增大,水贯穿石块的概率也会增大。所谓渗流相变(Percolation transition),即是当孔洞间存在通道的概率增大到某个临界值后,由水贯通的孔洞能够形成一个系统级(System level)的连通分支(例如,连通分支所包含的孔洞占总孔洞数的1%以上),而在此之前类似的系统级连通分支并不存在。例如图3(B)中的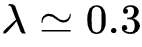 即是一个可能的相变点,在此之后一个系统级的强连通分支出现在功能连接中。这一新涌现出的系统级连通分支暗示着新属性出现的可能。
即是一个可能的相变点,在此之后一个系统级的强连通分支出现在功能连接中。这一新涌现出的系统级连通分支暗示着新属性出现的可能。
图3. 作为渗流过程的功能连接形成。(A) 大脑功能连接与多孔石块的类比。(B) 不同λ条件下功能连接中的最大强连通分支的情况。(C) 渗流相变点的求解过程(具体含义见论文正文推导)。(D) Non-dilution条件下渗流理论预测的一个神经元属于最大强连通分支的概率与图2结果中的真实概率的高度一致性。(E) Dilution 和 Non-dilution 条件下渗流分析的区别。(F)Dilution条件下渗流理论预测的渗流相变点与Non-dilution条件下渗流情况的一致性。
为了符合神经科学需求,作者区分了两类由λ控制的渗流过程:一种为对应在刺激输入后,任意神经元都有一定概率被突触前神经元激发的Non-dilution条件;一种为对应刺激输入后,神经元除了考虑和突触前神经元的并发外,还考虑对刺激的选择性(例如,神经元可能对某些刺激无响应,而对另一些神经元有强烈响应)的Dilution条件。
基于图3(C-F)的结果,作者发现无论是Dilution和Non-dilution条件下的渗流过程,都在果蝇大脑中对应着一个相变点(理论解和实验值高度匹配),这与图3(B)中朴素的观察一致。同时,在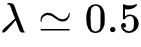 之后,功能连接中系统级的强连通分支的增大速度放缓,并在
之后,功能连接中系统级的强连通分支的增大速度放缓,并在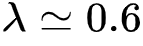 之后受到静态连接的约束而达到上界。
之后受到静态连接的约束而达到上界。
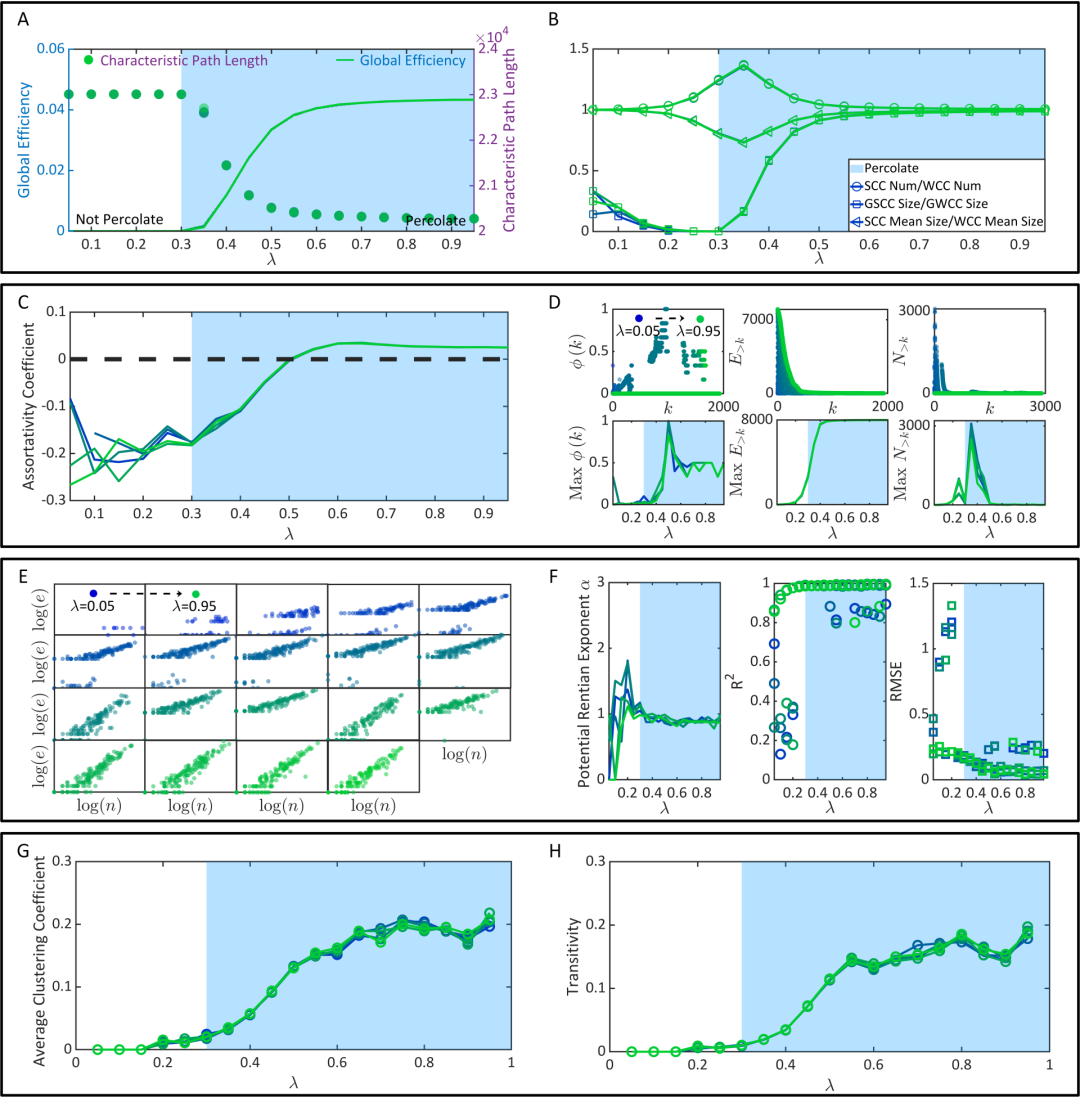
作者在对应不同的λ条件的功能连接上计算了衡量大脑的效率、鲁棒性和经济性的各类指标。这些指标由不同研究论文基于不同理论方法提出,并没有必然的关联性。然而,我们可以发现,上述指标(1)或者在渗流相变前(图4中白色背景区域)数值较低/较高,并在渗流相变后(图4中蓝色背景区域)数值迅速增大/降低,直到达到由静态连接决定的上界/下界;(2)或者在渗流相变前数值无规律剧烈抖动,在渗流相变后数值趋于收敛。
图4. 大脑效率、鲁棒性和经济性的涌现。(A-B) 用于衡量大脑信息传输效率的各类指标随着渗流过程的变化。(C-D) 用于衡量大脑鲁棒性的各类指标随着渗流过程的变化。(E-F) 用于衡量大脑在网络形成方面的经济性的各类指标随着渗流过程的变化。(G-H) 用于衡量大脑在网络运行方面的经济性的各类指标随着渗流过程的变化。
上述结果证明,大脑的效率、鲁棒性和经济性——无论其是用何种变量进行刻画——都随着时渗流相变的发生而迅速涌现,相应属性的涌现会在之后减慢,并在之后达到上界。考虑到之后相应属性的涌现速度已经明显放缓,不足以平衡λ继续增大所带来的额外能量消耗(兴奋性连接越多,神经动力学越剧烈,能耗越大),所以上述结果暗示是一个相对适合保持大脑优越属性的突触E/I balance。在论文中,作者还基于 [30] 提供的方法将预测的最优突触 E/I balance换算为对应的神经元E/I balance(兴奋性神经元占总神经元的比重),与实验中哺乳动物大脑中兴奋性神经元占比更高这一观测相一致 [31],且与其他关于E/I balance的理论和实验研究建立了联系。
总之,该研究结合细粒度的果蝇全脑连接组、大规模神经动力学计算和统计物理理论,发现了大脑效率、鲁棒性和经济性的涌现与由突触E/I balance调控的渗流过程的紧密关系,并基于此预测了最优的突触E/I balance,与实验观测相一致。在 [32] 中,作者开源了大规模神经动力学计算的数据以供后续研究者使用。
该研究由清华大学国强研究院人工智能与机器人领域自由探索研究项目 (2020GQG1017) 提供资助。作者感谢华为2012实验室中央研究院先进计算与存储实验室的Yaoyuan Wang和Ziyang Zhang研究员提供的支持,也特别致谢集智俱乐部的编辑在本文稿写作过程中提供的帮助。
[1] Mišić, B., Sporns, O., & McIntosh, A. R. (2014). Communication efficiency and congestion of signal traffic in large-scale brain networks. PLoS Computational Biology, 10(1), e1003427. https://doi.org/10.1371/journal.pcbi.1003427, PubMed: 24415931
[2] Avena-Koenigsberger, A., Misic, B., & Sporns, O. (2018). Communication dynamics in complex brain networks. Nature Reviews Neuroscience, 19(1), 17–33. https://doi.org/10.1038/nrn.2017 .149, PubMed: 29238085
[3] Amico, E., Abbas, K., Duong-Tran, D. A., Tipnis, U., Rajapandian, M., Chumin, E., … Goñi, J. (2021). Towards an information theoretical description of communication in brain networks. Network Neuroscience, 5(3), 646–665. https://doi.org/10.1162/netn _a_00185, PubMed: 34746621
[4] Graham, D., Avena-Koenigsberger, A., & Misic, B. (2020). Network communication in the brain. Network Neuroscience, 4(4), 976–979. https://doi.org/10.1162/netn_e_00167, PubMed: 33195944
[5] Avena-Koenigsberger, A., Misic, B., & Sporns, O. (2018). Communication dynamics in complex brain networks. Nature Reviews Neuroscience, 19(1), 17–33. https://doi.org/10.1038/nrn.2017 .149, PubMed: 29238085.
[6] Kaiser, M., Martin, R., Andras, P., & Young, M. P. (2007). Simulation of robustness against lesions of cortical networks. European Journal of Neuroscience, 25(10), 3185–3192. https://doi.org/10.1111 /j.1460-9568.2007.05574.x, PubMed: 17561832.
[7] Joyce, K. E., Hayasaka, S., & Laurienti, P. J. (2013). The human functional brain network demonstrates structural and dynamical resilience to targeted attack. PLoS Computational Biology, 9(1), e1002885. https://doi.org/10.1371/journal.pcbi.1002885, PubMed: 23358557
[8] Aerts, H., Fias, W., Caeyenberghs, K., & Marinazzo, D. (2016). Brain networks under attack: Robustness properties and the impact of lesions. Brain, 139(12), 3063–3083. https://doi.org/10 .1093/brain/aww194, PubMed: 27497487
[9] Avena-Koenigsberger, A., Mišić, B., Hawkins, R. X., Griffa, A., Hagmann, P., Goñi, J., & Sporns, O. (2017). Path ensembles and a tradeoff between communication efficiency and resilience in the human connectome. Brain Structure and Function, 222(1), 603–618. https://doi.org/10.1007/s00429-016-1238-5, PubMed: 27334341
[10] Kaiser, M., & Hilgetag, C. C. (2004). Edge vulnerability in neural and metabolic networks. Biological Cybernetics, 90(5), 311–317. https://doi.org/10.1007/s00422-004-0479-1, PubMed: 15221391
[11] Achard, S., Salvador, R., Whitcher, B., Suckling, J., & Bullmore, E. (2006). A resilient, low-frequency, small-world human brain functional network with highly connected association cortical hubs. Journal of Neuroscience, 26(1), 63–72. https://doi.org/10 .1523/JNEUROSCI.3874-05.2006, PubMed: 16399673.
[12] Alstott, J., Breakspear, M., Hagmann, P., Cammoun, L., & Sporns, O. (2009). Modeling the impact of lesions in the human brain. PLoS Computational Biology, 5(6), e1000408. https://doi.org/10 .1371/journal.pcbi.1000408, PubMed: 19521503
[13] Crossley, N. A., Mechelli, A., Scott, J., Carletti, F., Fox, P. T., McGuire, P., & Bullmore, E. T. (2014). The hubs of the human connectome are generally implicated in the anatomy of brain disorders. Brain, 137(8), 2382–2395. https://doi.org/10.1093 /brain/awu132, PubMed: 25057133.
[14] Bullmore, E., & Sporns, O. (2012). The economy of brain network organization. Nature Reviews Neuroscience, 13(5), 336–349. https://doi.org/10.1038/nrn3214, PubMed: 22498897
[15] Sporns, O. (2011). The non-random brain: Efficiency, economy, and complex dynamics. Frontiers in Computational Neuroscience, 5, 5. https://doi.org/10.3389/fncom.2011.00005, PubMed: 21369354.
[16] Hahn, A., Kranz, G. S., Sladky, R., Ganger, S., Windischberger, C., Kasper, S., & Lanzenberger, R. (2015). Individual diversity of functional brain network economy. Brain Connectivity, 5(3), 156–165. https://doi.org/10.1089/brain.2014.0306, PubMed: 25411715.
[17] Friston, K. (2010). The free-energy principle: A unified brain theory? Nature Reviews Neuroscience, 11(2), 127–138. https://doi.org/10 .1038/nrn2787, PubMed: 20068583.
[18] Laughlin, S. B., van Steveninck, R. R. d. R., & Anderson, J. C. (1998). The metabolic cost of neural information. Nature Neuroscience, 1(1), 36–41. https://doi.org/10.1038/236, PubMed: 10195106.
[19] Karbowski, J. (2007). Global and regional brain metabolic scaling and its functional consequences. BMC Biology, 5(1), 1–11. https://doi.org/10.1186/1741-7007-5-18, PubMed: 17488526.
[20] Betzel, R. F., Avena-Koenigsberger, A., Goñi, J., He, Y., De Reus, M. A., Griffa, A., … Sporns, O. (2016). Generative models of the human connectome. NeuroImage, 124, 1054–1064. https://doi .org/10.1016/j.neuroimage.2015.09.041, PubMed: 26427642.
[21] Kaiser, M., & Hilgetag, C. C. (2006). Nonoptimal component placement, but short processing paths, due to long-distance projections in neural systems. PLoS Computational Biology, 2(7), e95. https://doi.org/10.1371/journal.pcbi.0020095, PubMed: 16848638.
[22] Rubinov, M., Ypma, R. J., Watson, C., & Bullmore, E. T. (2015). Wiring cost and topological participation of the mouse brain connectome. Proceedings of the National Academy of Sciences, 112(32), 10032–10037. https://doi.org/10.1073/pnas.1420315112, PubMed: 26216962.
[23] Goñi, J., Van Den Heuvel, M. P., Avena-Koenigsberger, A., De Mendizabal, N. V., Betzel, R. F., Griffa, A., … Sporns, O. (2014). Resting-brain functional connectivity predicted by analytic measures of network communication. Proceedings of the National Academy of Sciences, 111(2), 833–838. https://doi.org /10.1073/pnas.1315529111, PubMed: 24379387.
[24] Hermundstad, A. M., Bassett, D. S., Brown, K. S., Aminoff, E. M., Clewett, D., Freeman, S., … Carlson, J. M. (2013). Structural foundations of resting-state and task-based functional connectivity in the human brain. Proceedings of the National Academy of Sciences, 110(15), 6169–6174. https://doi.org/10.1073/pnas .1219562110, PubMed: 23530246.
[25] Sporns, O., Tononi, G., & Kötter, R. (2005). The human connectome: A structural description of the human brain. PLoS Computational Biology, 1(4), e42. https://doi.org/10.1371/journal.pcbi .0010042, PubMed: 16201007.
[26] Xu, C. S., Januszewski, M., Lu, Z., Takemura, S.-Y., Hayworth, K. J., Huang, G., … Plaza, S. M. (2020). A connectome of the adult Drosophila central brain. BioRxiv. https://doi.org/10.1101/2020 .01.21.911859.
[27] Kaiser, M., & Hilgetag, C. C. (2004). Edge vulnerability in neural and metabolic networks. Biological Cybernetics, 90(5), 311–317. https://doi.org/10.1007/s00422-004-0479-1, PubMed: 15221391
[28] Breskin, I., Soriano, J., Moses, E., & Tlusty, T. (2006). Percolation in living neural networks. Physical Review Letters, 97(18), 188102. https://doi.org/10.1103/PhysRevLett.97.188102, PubMed: 17155581.
[29] Humphries, M. D., Gurney, K., & Prescott, T. J. (2006). The brainstem reticular formation is a small-world, not scale-free, network. Proceedings of the Royal Society B: Biological Sciences, 273(1585), 503–511. https://doi.org/10.1098/rspb.2005.3354, PubMed: 16615219.
[30] Agrawal, V., Cowley, A. B., Alfaori, Q., Larremore, D. B., Restrepo, J. G., & Shew, W. L. (2018). Robust entropy requires strong and balanced excitatory and inhibitory synapses. Chaos: An Interdisciplinary Journal of Nonlinear Science, 28(10), 103115. https:// doi.org/10.1063/1.5043429, PubMed: 30384653
[31] Sahara, S., Yanagawa, Y., O’Leary, D. D., & Stevens, C. F. (2012). The fraction of cortical gabaergic neurons is constant from near the start of cortical neurogenesis to adulthood. Journal of Neuroscience, 32(14), 4755–4761. https://doi.org/10.1523 /JNEUROSCI.6412-11.2012, PubMed: 22492031
[32] Tian, Y., & Sun, P. (2021). Co-activation probability between neurons in the largest brain connectome of the fruit fly. Zenodo. https://doi.org/10.5281/zenodo.5497516
随着电生理学、网络建模、机器学习、统计物理、类脑计算等多种技术方法的发展,我们对大脑神经元相互作用机理与连接机制,对意识、语言、情绪、记忆、社交等功能的认识逐渐深入,大脑复杂系统的谜底正在被揭开。为了促进神经科学、系统科学、计算机科学等领域研究者的交流合作,我们发起了【神经动力学模型读书会】。
集智俱乐部读书会是面向广大科研工作者的系列论文研读活动,其目的是共同深入学习探讨某个科学议题,激发科研灵感,促进科研合作。【神经动力学模型读书会】由集智俱乐部和天桥脑科学研究院联合发起,已于3月19日开始,每周六下午14:00-16:00(或每周五晚上19:00-21:00,根据实际情况调整)进行,预计持续10-12周。期间将围绕神经网络多尺度建模及其在脑疾病、脑认知方面的应用进行研讨。
详情请见:
神经动力学模型读书会启动:整合计算神经科学的多学科方法
点击“阅读原文”,报名读书会