脑机类比特刊:寻找复杂智能系统的普遍规律

导语
大脑是已知最为复杂的系统,而构建类脑的人工智能机器是人类科技的高远目标。但怎样利用对大脑结构与功能的认识,进而构建通用人工智能,是多个学科领域的长期目标。本篇综述以「脑机类比的当前思考——所有的隐喻都是错误的,但有些是有用的」为研究主题,对收录在 Frontiers Ecology and Evolution 特刊中的 12 篇文章进行了详细梳理。
为便于读者理解,本文用矩阵的方式将选定的概念与对应的文章联系起来(见图 1)。本期特刊的所有作者一致认为语义学(semantics)是脑机类比辩论中的一个关键问题,并且对这些问题进行了深入研究。但是,目前的研究缺乏对隐喻(metaphor)和类比(analogy)之间区别的认识。为此,本文重新评估并描述了这一区别,并为后者提供了定义。此外,作者们还认为有必要发展大脑的进化理论,并为此提供了一个概述。文章以对科学创造力的思考作为结尾,认为:无论是在科学还是艺术领域,隐喻和类比及其美学影响对于创造过程都至关重要。

Giorgio Matassi,Pedro Martinez | 作者
贾圣祎 | 译者
朱欣怡 | 审校
邓一雪 | 编辑
论文题目: The brain-computer analogy—“A special issue”
论文地址:
https://www.frontiersin.org/articles/10.3389/fevo.2022.1099253/full
1. 导言
2. 特刊论文简介
3. 讨论延伸
1. 导言
1. 导言
哲学家、心理学家、数学家、物理学家、计算机科学家和神经科学家一直在比较大脑和计算机。有些作者认为这是一种空洞的讨论(他们认为大脑“显然是”计算机),而也有观点认为,这两个系统的运作中都有一些实例不能轻易得出结论,需要进一步分析。研究者对脑机类比(Computer Analogy)的困惑来源于以下事实:
第一,许多研究人员不理解智能行为不等于信息处理器(计算机似乎利用处理器表现出智能行为,并不意味着「智能」和「信息处理」是等同的,这是2016 年,心理学家 Robert Epstein 在《The empty brain》中表达的观点);
第二,一些人认为给大脑分配一个诸如「计算系统」的名称是有局限性的,而且会扭曲我们对大脑过程的观察,如意识和感知。
在这种情况下,我们认为重新探讨脑机类比仍然是非常有意义的。事实上,本期特刊所收录的 12 篇讨论脑机类比的文章都表明这个主题(或争论)仍然非常活跃。
1.1 研究课题
从一开始,我们就清楚地看到,该项目将以网络结构进行组织,因为其主要概念在不同程度上相互关联。我们推断,网络结构将有利于「信息」在节点(学科、方法、文章)之间交换。通过这种组织,我们能够将研究对象(大脑)和研究方法视为一个相似现象的两种表现形式,而节点(思想、方法或物理实体)的网状互联则是这些连接的图形化表达。
一系列作为「网络节点」的主题和概念构成了该项目的基础:概念点(哲学)、网络科学、复杂系统(自组织)、神经网络与计算神经科学(人工神经网络)、计算机科学(分布式-集中式体系结构;丘奇-图灵论题;计算复杂度)、信息论(可靠性-误差检验;信息的效率与速度;信息不对称)、博弈论(分布式神经结构;信息分布不对称)、量子脑-量子计算机、人工智能(AI)与人工生命、理论神经科学、大脑进化(进化发育)。
该领域的历史发展告诉我们,来自网络科学的知识(如 A-L. Barabasi、MEJ Newman、DJ Watts 等人的工作)有助于理解大脑如何工作、互动、灵活地管理任务,以及突触分布、密度和强度如何潜在地影响这些活动。网络进化可以阐明神经网络结构的进化历史(及其控制原理;Sterling 和 Laughlin,2017)。研究者们运用不同的方法(细胞神经科学、计算建模、连接组学分析、神经科学哲学等)研究网络进化,并对网络在大脑功能中的角色提出了不同的观点。其中,大多数研究处理一般的(大脑网络的)表示问题,而新近的研究则将目光移到了信息流(路由)上。如在本期特刊的一篇论文中,D. Graham 提出互联网的功能模式是理解大脑功能的一个新参考框架。
在本期特刊中,我们希望解决的另一个与网络进化相关的重要问题是:自组织和复杂系统在塑造大脑架构及其进化中的作用(如 I. Prigogine and G. Nicolis, C. G. Langton, S. Kauffman, S. Kelso, P. Bak, S. H. Strogatz, C. Gershenson and F. Heylighen, R. Solé等人的工作)。“涌现属性”(emergent properties)或“组织层次”等概念在这里也是相关的。
最后,我们很清楚大脑和计算机之间的关系涵盖了自然科学、数学、计算机科学、心理学和哲学等一系列广泛的主题,因此我们需要缩小研究范围。我们选择不涉及意识、行为、语言和文化等领域。
在讨论本期特刊中的文章(见表 1)以及它们对辩论的贡献之前,我们回顾一些关键且必要的概念/主题:机器、科学中的隐喻和类比、大脑。在下文中,属于本特刊的参考文献以出版年份后的(*)标识。
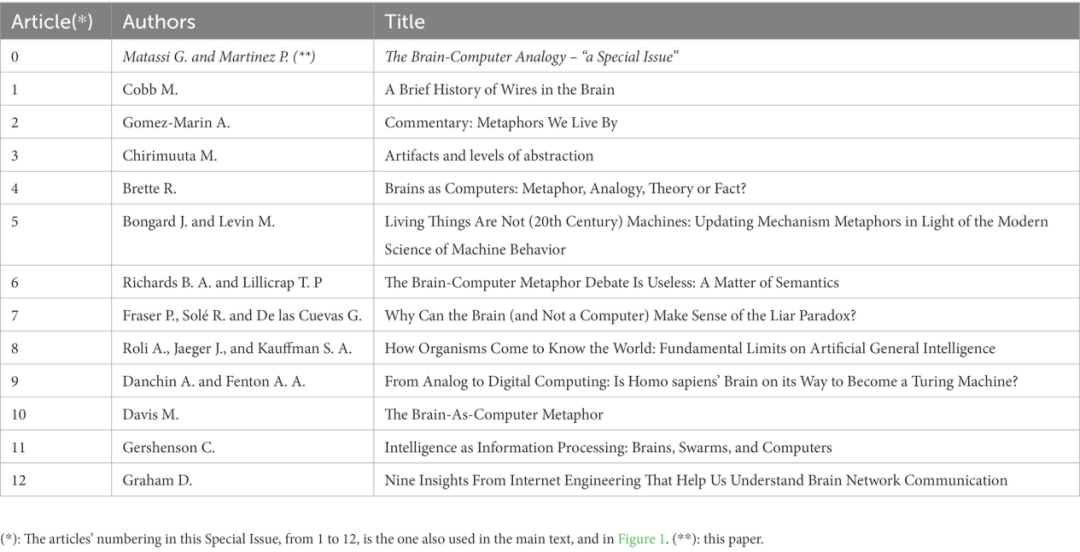
表1.特刊中的文章。
1.2 语义学:概念和定义
「我提议考虑这样一个问题:『机器能思考吗?』这应该从定义『机器』和『思考』这两个术语的含义开始。」(图灵,1950)毫无疑问,艾伦·图灵清楚地认识到语义学在这方面的重要性。同样,本期特刊的所有作者和编辑都认为语义学(semantics)是脑机类比辩论中的一个关键问题。
作者们认为有一些术语的当前定义存在问题,包括「计算机」、「计算」、「递归」、「算法」、「可计算函数」、「机器人」、「程序」、「软件」、「信息」、「人工智能」、「智力」、「认知」。
在下文中,我们将重点讨论两个具体的基本问题:「机器」的定义以及「隐喻」与「类比」的区别。对于前者,在本期特刊中,Cobb 主要处理历史上大脑的图像,Bongard 和 Levin 表达了他们对术语「机器」的一种「过时」观点的担忧。在本文中,我们将追溯「不同种类的机器」是如何最好地代表大脑的历史。至于后者,只有 Brette 和 Gomez-Marin 才涉及到隐喻和类比的区别。因此,我们认为有必要对这些概念进行更详细的描述,并为后者提供一个修订的定义。
1.3 机器
经过几十年的研究,神经科学家们形成了一个普遍的观点,即大脑「是什么」取决于你如何研究它。我们生活在一个机械时代,所以我们把大脑当作一台机器来研究。在这种背景下,我们应该首先问自己:机器是什么?我们对机器的看法如何随着时间的推移而改变?在本期特刊中,Cobb和Bongard/Levin介绍了我们过去和现在理解机器的方式。
长期以来,大脑被同化为某种「机器」。这一思想可以追溯到笛卡尔哲学,把机器理解为能够执行某些功能的任何物理系统。笛卡尔认为:身体器官以纯粹机械的方式运作,他「创造性地」改编了以前(亚里士多德、盖伦等)的理论。
多年来,机器类比的形式经历了许多转变,但机械世界观(液压、电气或信息)一直占主要地位。在本期特刊中,Cobb和Bongard/Levin分别回顾了一些历史上的和人工智能最新进展的观点。有趣的是,随着20世纪分子生物学的进步,机器类比的视域已经从整个组织转向控制其不同功能的生化成分。从这个意义上讲,大脑等同于由高度协调的化学成分(分子机器)组成的一锅汤,这些化学成分使我们获得丰富的心理体验。在18、19世纪之交从欧洲(尤其是德、法两国)萌芽、并在20世纪六、七十年代迅猛发展的神经化学,认为关联神经系统中的化学和功能是理解大脑的最有效途径。
从功能的角度来看,我们对大脑的看法经历了从「被动的流体传导装置」到「主动的信息处理装置」的转变。计算机(原意是一个能够进行「计算」操作的人)一直被理解为具有某些特性(存储、检索和处理数据的能力)的机械设备。在这个阶段,我们应该考虑到,尽管英语单词「computer」的意思是表示可以存储、检索和处理数据的可编程机器(大英百科全书),但其他的一些罗曼语族(Romanic languages)的语言保留了计算机作为组织或计算数据集的人的原始含义(例如,法语:ordinateur和西班牙语:ordenador)。现在,我们使用计算机这个术语表示一种根据一组指令来处理数据的设备(通常是电子设备),但值得记住的是Mahoney在他对计算(computing)的历史回顾中(Mahoney,1988)给出了计算机更精确的定义:一个基本上由三部分组成的结构。这反映了与这种「机器/设备」的性质有关的三个历史学科的贡献(电气工程、计算机科学和软件工程)。此外,Mahoney在总结句中明确说明了这些贡献:「数学奠定理论基础、电子学使其实际可行、编程使其具有智力并让它融入到经济社会当中」。
随着信息时代的到来(20世纪40年代),信息内容和逻辑运算被阿隆佐·邱奇(Alonzo Church)和艾伦·图灵(Alan Turing)等逻辑学家引入,这时对大脑结构和功能最突出的类比是计算机,即所谓图灵机(TM)的实例。焦点已经从机器的实例化(instantiation )变为底层操作。图灵在他1950年的论文中将「机器」描述为「……每一种工程技术」制造的人工制品,并建议将它们与「电子计算机」或「数字计算机」等同起来,因为他对这些设备的历史感兴趣(图灵,1950)。他把计算机定义为有限状态机(一个计算的数学模型)。这里引用图灵的一句话似乎是恰当的:
“一台数字计算机通常可被视为由三部分组成:(一)存储[信息]……相当于人类计算机(human computer)使用的纸张……[和]……规则书”,(二)执行单元[进行计算],(三)控制[正确使用规则]。…数字计算机…属于离散状态机(discrete state machines)的范畴。…这种机器可以抽象地描述如下。机器的内部状态(由轮子的位置描述)可能是q1、q2或q3。有一个输入信号i0或i1(杠杆的位置)。根据[规则]表,任何时刻的内部状态都由最后的状态和输入信号决定。……这些机器通过突然的跳跃或点击从一个非常确定的状态移动到另一个状态。……数字计算机……必须为它想要模仿的每个新机器重新编程。数字计算机的这一特殊属性……可以概括为通用性”。
当然,并不是所有的脑机隐喻都依靠传统的图灵机或冯·诺依曼架构。例如,并行计算或量子计算等模式丰富了我们对计算机能做什么的看法。然而,关于计算和大脑功能的一个持久观点强调了两者都利用的并行架构,将问题分解成更小单元并由不同组件处理,所有组件通过共享内存进行通信。因此,比较计算机和大脑时,一个共同的推论是:这两个系统本质上都依赖于并行处理器。这不是相似性的准确表示,原因有很多:(一)大脑和计算机的独立(计算)单元的数目具有6到7数量级的差异。(二)虽然计算机中的处理器是「通用的」,但人类大脑有专门处理不同类型输入的特定区域。(三)大脑和计算机之间在可靠性和适应性方面存在很大差异(这一概念与两个系统中的「可重新编程性」相关),其中大脑信息处理系统本质上是「噪声(noisy)」的。这些因素似乎表明,大脑中不存在真正的并行处理,大脑和计算机中的重新编程依赖于不同的网络「重新配置」策略(机器中的重新路由与生命系统中的神经可塑性)。生物系统的适应性(例如,脑)是从细胞和回路结构的可塑性得到的独特性质,并且是控制脑神经元集合的出生、死亡和相互连接的遗传(genetic)和表观遗传(译者注:epigenetic,指在「非DNA序列变化」情况下,遗传信息通过某些机制或途径,发生可保存并传递给子代的基因表达或细胞表型之改变)因素的结果。
在下文中,我们返回到处理系统并且提供图灵机(TM)的「无障碍」描述,其应当有助于理解用于大脑的隐喻。简单地说,图灵机,或者图灵所称的「自动机器」(图灵,1937),是数字计算机的一种抽象的、理想化的模型。机器的输入是一串符号,每一个符号由线性磁带上的一个单元携带。这台机器拥有某种读写扫描头,每次只处理一个单元。它是自动机器(即,在任何给定时刻,其行为完全由当前状态和符号、以及被扫描的「配置(configuration)」确定)。它是一台能够进行有限组配置的机器。有限的规则集(即,表示算法的程序)指示机器如何响应每个符号(即:擦除、写入、向左移动、向右移动、不移动)。原则上,对于任何可计算(computable)的函数(即其值可以通过算法来计算的函数),都存在能够计算它的图灵机。经逻辑推理我们可得出,一个通用图灵机(UTM)能够通过从磁带上读取相应的规则集,模拟任何图灵机执行不同的任务。这就是可编程计算机(programmable computer)的理论模型。
给出图灵机的定义后,很快就可以清楚地看到,大脑(或心智)可以等同于一个与图灵机非常相似的计算系统,并且许多心理过程与图灵机执行的计算非常相似。一些作者认为,这种将大脑与图灵机等同起来的立场非常强硬,因此被称为「硬立场(hard position)」。
这一「硬立场」受到了其他作者的批评,批评者认为,无论是从大脑的物质组成和还是运行原理上来讲,大脑都不能等同于图灵机(除了两者进行算术运算的方式)。我们不打算在这里深入探讨这个问题,只想强调图灵机在过去80年里所产生的巨大影响,正如计算神经科学家(并非所有)所坚持的那样,他们认为大脑就是计算机(作为图灵工作)。当神经科学家们意识到神经元在以「全有或全无」的方式执行其生理功能,即激发动作电位时,这个模型就得到了证实。1943年,Warren McCulloch和Walter Pitts认为大脑中的神经回路是逻辑门电路。现代神经科学揭示了更复杂的放电模式和放电调控模式,为最初的麦卡洛克-皮茨观点增加了细微的差别。
在比较计算机和大脑时,语义问题常常以这样或那样的形式被提出来。特别是,在80年代约翰·塞尔(John Searle)问的问题:「一台机器真的能被称为智能吗?」(Searle,1984)。这一问题包含在著名的「中文房间(Chinese room)」思想实验中。这个实验表明,不管你怎样编写计算机程序,计算机还是不懂中文;它只是模仿知识,因此这种行为不能等同于智力。塞尔认为,他的思想实验强调了这样一个事实,即计算机仅仅使用句法规则来操纵符号序列,而没有理解它们的含义。「含义(meaning)」的问题在这里没有进一步探讨,尽管我们认可它的研究价值。塞尔的主要结论是,通过图灵测试并不足以作为他所提出问题的答案。
总而言之,尽管脑-机类比在历史上有过好运,也有过不幸,但这种「隐喻」的使用还是很普遍的,这一点的证明可以在本期特刊的不同论文中找到。这个问题仍然很有价值,可以进一步讨论。
1.4 科学中的隐喻与类比
关于隐喻(metaphor)和类比(analogy)的定义,尽管在科学和人文学科中有大量的文献致力于研究它们,但这些概念仍然是混乱的,这一点去看看他们在英语词典和百科全书中的定义就能发现。一个生动的例子来自韦氏大词典(Merriam-Webster Collegiate Dictionary),其中隐喻被定义为「一种修辞方式,用一个词或短语表示一种物体或想法,以暗示它们之间的相似性或类比。」
这两个概念的定义在人类知识库的不同分支中得到了「调整」。这里我们只讨论(西方)科学思想中隐喻和类比的含义。我们将在以下章节中看到,在科学界和本期特刊中,隐喻和类比被当作同义词,尽管它们存在差别。正因如此,我们在以《脑机类比的当前思考——所有的隐喻都是错误的,但有些是有用的》的研究课题中,有意且挑衅性地将隐喻和类比作为同义词使用;而正是在科学中,两个词的区别更为明显。这就是本节详细讨论这一问题的理由。
值得注意的是,一些人认为隐喻和类比对于科学是无足轻重的,另一些人则声称隐喻和类比对科学创造力至关重要(本文支持后者)。根据Ziman的说法,「……科学理论不可避免地带有隐喻性」(Ziman,2000),有人认为它们是「人类扩展自身知识边界的能力的基础」(Yohan,2012)。此外,隐喻和类比推理的能力是人类认知的重要组成部分。不可否认,隐喻和类比是传播知识的一种强有力的方式,因而也是教育和学习的一种有力工具。想想在我们自己的教学经验中,我们有多少次使用隐喻向学生传达概念(Kovac,2003)。隐喻和类比可以指导科学发现、假设和理论,并且在使世界理解科学语言方面也发挥着重要作用。正如Kuhn所说“隐喻在建立科学语言和世界之间的联系方面起着至关重要的作用”,但关键的是,”……然而,这些联系不是一劳永逸的”(Kuhn,1993)。那么,选择“正确的”隐喻可被视为科学家/教师工作的一部分,最终成为一种艺术形式(Haack,2019)。鉴于我们对大脑的讨论与隐喻和类比关系密切,在下文中,我们将更详细地讨论它们。
1.5 Metaphor 隐喻
关于隐喻的文献铺天盖地,定义也很多。Robert R. Hoffman 认为科学隐喻以各种不同的形式出现,具有各种不同的功能,并列出了相当详尽的清单(Hoffman,1985)。一个例子是“知识树(Tree of Knowledge)”。几个世纪以来,知识树有着各种各样的风格,它们超出了科学范畴,可以说是人类文明的基础隐喻(Lima,2014)。而且,对于进化生物学家(如本文的两位作者)来说,几乎没有比生命树更基本的隐喻了,达尔文和他之前的拉马克(Lamack,1809)曾用它来说明进化论,并在“分化分类群图”(Diagram of diverging taxa,Darwin,1859)中进行了描述(第6版中的116 – 117)。事实上,树的比喻一直被用于进化生物学。然而,值得注意的是,在不到三十年前,基于对物种间基因流动现象的进化影响的重新认识(又名:水平/垂直基因转移 Horizontal/Lateral Gene Transfer),一个解释物种的多样性的新隐喻出现了:生命网络(The Network of Life)。顺便提到的是,事实上,网络隐喻的提出早于生命树的隐喻。前者在1750年由Vitaliano Donati提出,后者则在1776年由Pallas提出。在进化生物学中使用树和网络的例子在这里被特别提及,以强调这两种隐喻的互补性;我们认为这一立场在本综述中是关键的,因为树和网络都被用于模拟我们对大脑及其进化的想法(见第3节)。
至于科学中隐喻的定义,为简洁起见,我们挑出其中两个(JC Maxwell, 以及Lakoff & Johnson),并辅以三个说明性的例子,以便更好地理解。
詹姆斯·克拉克·麦克斯韦(James Clerk Maxwell)写道:「我们把一门熟悉的科学的语言和思想迁移(transfer)到一门我们不太熟悉的科学中去的修辞或思维方式,可以称为科学隐喻。」这指的是隐喻的「逻辑语义学」观点,在科学和日常生活中大量使用。
然而,在Lakoff和Johnson的经典定义中,也引入了「映射(mapping)」的概念。他们说:「隐喻的本质是用一种事物来理解和体验另一种事物」(Lakoff和Johnson,1980),同时也引入了不同的观点,即「隐喻是我们理解抽象概念和进行抽象推理的主要机制……隐喻是跨概念域(conceptual domains)的映射」(Lakoff,1993a)。因此,隐喻「成为」概念工具(以帮助理解)。正如Humar所说:「隐喻通过将属性从一个映射到另一个来连接两个域。因此,隐喻是一种转移行为…… [其中] …… Lakoff和Johnson引入了关键术语『目标(target)』和『来源(source)』……例如,生物学隐喻『基因是文本』将来源『文本』和目标『基因』联系起来」(Humar,2021)。Black(1962)指出,在这种情况下,《牛津英语词典》对隐喻的“标准化”描述与上述描述是多么的相似:「(隐喻是)一种修辞方式,将一个名称或描述性词语转移至与其本来适用的对象不同但类似的另一个对象;这种修辞方式的一个例子就是隐喻表达。」
有趣的是,「metaphora」一词的词源源于古希腊名词「metaphora」(μεταφορά),而「metaphora」又是由动词「metaphere」(μεταφέρειν)衍生而来,原意是「转移(transfer)」、「转化(transform)」,或由μετα(超过 over,超越 beyond)和πηερειν(携带 to carry)衍生。这完全取决于我们在上述定义中「转移」或「携带超越」的含义。更确切地说,我们可以问:转移的是什么?
在深入研究下一个相关的类比概念之前,我们需要考虑另一个经常与隐喻联系在一起或被比作隐喻的相当有问题的术语:「模型(model)」的概念。通常,在科学文献中,模型和隐喻之间没有明确的区别。但我们认为,为了明确起见,需要作出区分。与隐喻相反,模型(概念模型)的语义范围较窄,它作为系统的假设性表示,其目的是模拟和理解现实(例如,生物模型)。而且,「……一个模型,在其词源和技术意义上,是一个实质性的东西,它是其他东西的最好或理想的代表。「模型」一词的所有其他用法都是这一基本含义的隐喻延伸」(Hoffman,1985)。因此,我们将模型视为旨在理解和预测特定模式(pattern)的方法或表示。
在本节的末尾,我们希望提出一种隐喻「概念」,该概念不需要但接受映射概念的使用(参见下文1.6节)。我们脑海中的隐喻类型是建立在视觉感知的基础上的。视觉图像是科学洞察力的驱动力,并提供教育力量。作为一个例子,我们确定了三个最能说明这一思想的隐喻:Wright(1931)的《适应性景观(Adaptive Landscapes)》、沃丁顿Waddington(1957)的《表观遗传景观(Epigenetic Landscapes)》以及Davidson and Peter(2015)的《基因调控网络(Gene Regulatory Network )》。关于科学中隐喻的使用及其危险和陷阱,我们可以参考Reynolds(2022)的优秀新书。
1.6 类比
隐喻可能是「类比」(和「相似性」)的来源,并可能指导模型的构建。在《牛津英语词典》给出的类比定义中,包括以下内容:(a)比较两件事物,通常是为了解释或澄清;(b)生物学:具有不同进化起源的器官之间的功能相似性。我们在这里的重点是第一种,更一般的定义。
Atran(1990)将这个概念追溯到亚里士多德,他试图比较人、动物和植物的结构和功能;「… 亚里士多德的生命形式是通过拥有具有同样基本功能的类似器官来区分和联系的。」类似地,类比概念在比较解剖学中于1843年被引入,并被定义为「一种动物中的某个部位或器官与另一种动物中的另一个部位或器官具有相同功能」(Owen,1843)。Atran 提出类比概念的更一般化版本,并提到了牛顿的「自然类比」概念(同上,第232页),指出这个类比「…结合了两个旧观念:神学的『存在链(Chain of Being)』,自然(Nature)通过它寻求神的完美,以及宏观世界和微观世界中因果模式的统一。」
然而,正是在Mary Hesse(1970)详细描述科学模型和类比的经典著作中,她想象了两位科学家之间的对话:Campbellian 认为类比和「某种意义上的模型对于科学理论的逻辑是必不可少的」,而Duhemist则否认这一点。Campbellian 者将类比分为三种类型:正向、负向和中性。两个物理对象或系统具有正向类比,是基于它们共同的「属性」:「例如,地球和月球。两者都是大的、坚实的、不透明的、球形的物体,从太阳接收热和光,…」然而,这些物体在许多方面可能存在差异:「另一方面,月球比地球小,更具火山性质,没有大气层和水…它们之间存在负向类比。」 中性类比是「模型的属性,我们尚不确定它们是正向类比还是负向类比。」请注意,Campbellian 也关注语义:「但首先让我们就「模型」这个词的含义达成一致。」因此,Hesse 告诉我们,类比可以具有特定的「价值(value)」:正向、负向或中性。
Humar提出了「结构隐喻」和「功能隐喻」的二分法 (dichotomy)。实际上,「功能隐喻强调源语和目标语之间的功能相似性,在古代科学文献中也有类似的表述」(2021)。Gentner和Jezioreski(1993)也认为,任何类比概念的使用都贯穿着一个基本思想:「类比的核心思想是将知识从一个领域(基础,the base)映射到另一个领域(目标,the target),使得在基础对象之间存在的关系系统也存在于目标对象之间。在解释类比时,人们试图将基础对象与目标对象一一对应,以获得最大的结构匹配。」此外,Hoffman将这两个概念的区别视为鸡生蛋问题,将类比视为「心理的蛋」,将隐喻视为「鸡」(Hoffman,1985,第348页)。
最后,我们建议在科学中使用两种判断标准(结构和功能)定义类比,并将功能指定为其最具特色的属性。这样一来,我们所定义的类比就能与Lakoff-Johnson的隐喻定义以及其相关的「转移」行为联系起来。我们认为这个类比的定义在这里更为相关(具有实际意义),因为隐喻并不旨在为特定问题提供解决方案,它们没有解释能力。相反,类比确实具有解释能力,并使我们能够根据通过建立连接,基于从另一个系统所获得的知识理解给定系统的结构/功能。一个说明类比「解释作用」的典型例子是在不同学科之间转移的主题(Ziman,2000)的例子,例如从信息论到分子遗传学的「代码(code)」概念。
1.7 大脑
本特刊的一个关键概念显然是「大脑」,但如何定义大脑呢?根据《大英百科全书(Encyclopedia Britannica)》的简单定义,大脑是「生物体前端的神经组织团块」。大脑整合感官信息并指导运动反应。虽然这主要代表了脊椎动物的情况,但用神经元代替神经仍然是一个有效的论断。作为集中结构的大脑是古老的,可以追溯到埃迪卡拉纪(Ediacaran,5.71亿至5.39亿年前)的两侧对称动物的起源(Martinez和Sprecher,2020)。在幼虫/动物的一个极点中神经元的聚合,可以实现更好的集中功能协调,从这个意义上说,大脑也被等同于「中央处理单元」(CPU)。大脑是如何实现中央化的以及促使它们出现的条件已经被讨论过(同上),这里不需要再讨论。
几个世纪以来,我们对大脑的理解发生了彻底的变化,主要是由于对它们的物理组成部分和工作方式缺乏正确的理解。各种解释使用了符合所处时代机械世界的隐喻(见Cobb在此特刊中的历史梳理)。最近,随着计算设备的实例化和信息时代的崛起,计算和信息处理成为我们思考大脑及其活动的参考标的。目前的观点始于20世纪初,当时人们在显微镜下系统地分析了大脑组织。孤立的细胞形成的神经网络支持了将大脑视为一台「机器」来计算和处理信息的观点。
大脑连接(神经元和亚结构)的复杂性表明了大脑实际上是一组连通的电线,具有复杂的架构(Cobb,2020)。此外,化学和电学连接的发现树立了大脑作为一个巨大的电气设备的形象,它具有复杂的开关机制。正是信息时代中出现的第一批能够「计算」的设备才导致了大脑新模型的提出。大脑除了进行电脉冲传导外,还被同化(assimilate)为一个复杂的计算设备。
神经元的综合模型(integrative model)是由控制论/神经生理学先驱 Warren McCulloch和数学家Walter Pitts(以及其他人)发展的。McCulloch的控制论使用逻辑和数学来开发神经网络模型,以体现大脑的功能(Pitts and McCulloch, 1947)。模型的准确性成为辩论的主题,本期特刊中的一些论文也提到了这个问题(例如,Davis,2021*;Fraser,2021年*)。关于信息如何被处理、神经元通信的速度、神经元在整合输入方面的作用、信息的路由以及神经元发放模式和大脑活动(即心理活动)之间的相关性的思想,都为关于使用「计算机」隐喻来理解神经科学的不同方面的有效性的辩论做出了贡献。这个辩论今天仍然像过去一样活跃。
器官的活动作为特性「定义」了隐喻的范围和内容,这是使用特定器官系统的隐喻的基础。从这个意义上讲,大脑被比作计算机,因为根据某些作者的说法,它们实际上正在执行「计算」操作(Chirimuuta,2022*)。然而,对于该隐喻的使用,并没有形成统一意见(在Gomez-Marin和Graham的本期特刊中的文章也探讨了其他隐喻),这已经引起了关于在神经科学中使用某些特定隐喻的意义性的激烈辩论(请参见本文的后续讨论)。
对大脑及其功能进行建模的关键问题,与信息流的本质以及输入信号如何转化为输出行为(包括路由问题)相关(Graham, 2022*)。这和大脑与线性处理器的工作方式不同的观点有关。在这种情况下,从输入数据到输出实现(行为)的信息流并不是单向的。另有观点认为,大脑的输出是某些「涌现性质」的结果(emergent properties,译注:涌现性质是指在许多组分合作下出现的新的、高层次的物理化学属性或现象,而这些属性或现象是组分个体不具备的),这些涌现性质并不能由原始输入通过线性变换得出。不同的大脑物理模型无法解决的一些问题,与人类大脑的自指(self-reference)能力,或更一般地说,我们对自己存在的意识有关。这些问题不容易被涌现模型解决。在本期特刊中,Fraser 等人展示了自指模型在计算机科学中的复杂性。再次强调,数学上的描述和可观测的现实并不容易进行比较。
2. 特刊论文简介
2. 特刊论文简介
在本节中,我们对本期特刊中的12篇文章(见表1)进行了总结,其中除一篇外,其他每篇文章都得到了通讯作者的认可。
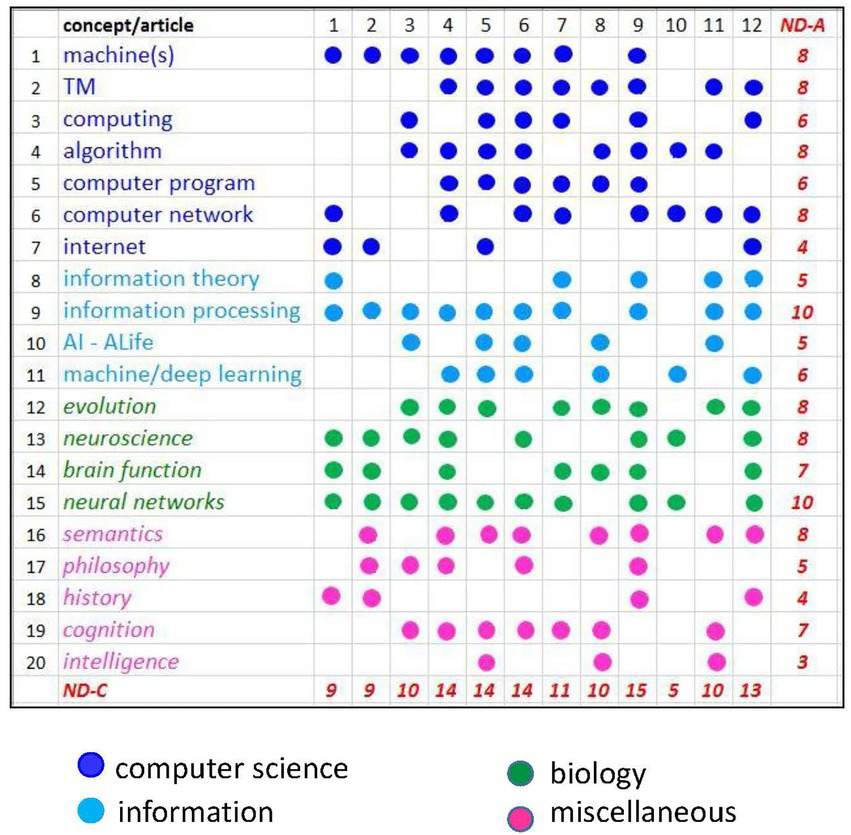
我们在图1中展示了一幅图片,可以看作是整个期刊的一个快照。它基于我们任意选择的20个概念(关键词),用[0,1]矩阵的方式表现论文中涉及的概念。这种表示方法的有用性不言自明。
我们选择Matthew Cobb的文章作为本期特刊的开篇,他对几个世纪以来用来描述大脑并试图理解其功能的隐喻进行了梳理。
2.1 脑电路简史
在本期特刊的开篇文章中,优秀著作《The Idea of the Brain》(2020)的作者Matthew Cobb详细介绍了大脑「线路图」(wiring diagram)隐喻的发展史,并探讨了它与相关隐喻在过去两个世纪对大脑概念的影响。
Cobb 认为使用隐喻来描述脑功能的记载始于18世纪,源于力学以及电学(电报)的发现。他认为,到了19世纪末期,「电话交换机的出现,信息得以灵活路由」,这是一个明显的转变。到了20世纪,「动物体和计算机的两种线路图形成了对话关系」。
在21世纪,旨在完整描述中枢神经系统结构连接的连接组(connectomic)项目成为了焦点。Cobb主要批评这些方法,因为它们产生了神经系统的静态表示。他认为,我们应该从控制特定行为的小型回路开始,逐步推进到整个神经元连接图谱。他以龙虾的胃为例,这个过程由几个神经元控制,且已经被研究了很长时间,但我们仍然没有完全理解其机制。此外,Cobb 指出,仅凭基因组序列本身无法解释相应有机体的「功能」,同样「……线路图本身也无法解释人类大脑的运作」。
Cobb 提到了被认为是最新的一种大脑功能隐喻的「云计算或互联网」。一方面,他承认「……它将可塑性和分布式功能融入了我们对大脑的概念中」,另一方面,他指出,在考虑到鲁棒性(robustness)概念时,它也有其局限性:「互联网被设计成即使移除了关键部分仍然能够运行,而大脑功能的某些方面如果特定区域受损则会受到决定性的破坏。」有趣的是,这种互联网隐喻将在David Graham在本特刊中的最后一篇文章中得到详细探讨。
作为一个警示,Cobb 警告我们使用这些隐喻研究大脑功能的局限性,主要是「……由于大多数神经系统的可塑性和分布式功能。」大脑可塑性的概念是本专题的核心,暗示了个体在其生命中表现出的神经系统可塑性与学习和记忆获取过程有关。
接下来的三篇文章涉及概念问题,由A. Gomez-Marin、M. Chirimuuta和R. Brette撰写。他们解决的问题是是否有任何基础来比较大脑和计算机。他们以不同的方式通过质疑计算机是什么、如何加以描述以及这些描述及其相关隐喻在我们对大脑和计算机的当前理解中的限制来回答这个问题。
2.2 评论:我们赖以生存的隐喻
Gomez-Marin向我们介绍了认知科学家George P. Lakoff和Mark Johnson所著的著名书籍《我们赖以生存的隐喻(Metaphors we live by)》(1980)。在这部开创性的作品中,作者对隐喻的本质进行了详细的分析,认为隐喻曾经被视为仅仅是「语言工具」(语义),而实际上它们是大多数「概念建构」,能够塑造我们的思维和行为方式。从某种意义上说,正如Gomez Marin所指出的,隐喻的语义角色是次要的,而它们的概念 (认知) 本质则更为重要。根据Lakoff (1993b) 的说法,隐喻映射(从一个概念领域到另一个概念领域)的进行独立于它们的语言表达,因此要优先考虑它们的认知功能,而不是语言表达的结构。换句话说,隐喻的概念结构比语言结构更为重要。
在这个背景下,Gomez-Marin重新审视了计算机和大脑的比喻。经过对大脑和计算机最普遍的视觉形象进行简要的历史回顾,Gomez-Marin让我们关注到例如全息图和收音机等较少被提及的大脑形象。其中,「收音机」的形象代表着一个有趣的可能性,即「大脑不会创造思想,只能接收和过滤它们。」
在最后,Gomez-Marin总结了他对隐喻的评价,并建议我们将它们作为实用工具应用,但需要时刻保持警惕,避免陷入「隐喻单一文化(metaphorical monoculture)」,否则将成为「负担(burden)」,而非「祝福(blessing)」。
2.3 组件与抽象水平
Chirimuuta评论了不同作者的观点,即多层次方法可以使大脑和计算机(或其他复杂工具)变得易于处理。这些方法使用自上而下的系统功能特征来补充自下而上的还原主义(reductionist)策略。重要的假设是:大脑可被分解为相对自治的组织层次,类似于计算机可被分解为硬件和软件。
图2. 逆向工程表示为功能分析的执行示意图
然而,尽管这个比喻很有吸引力,Chirimuuta认为还需要考虑一些限制因素。(1)底层组件(脑中的神经元)在脑信息处理中不仅仅是“硬件实现者”。在计算机中,维护机器物理完整性和执行信息处理任务的元素是不同的。而这种分离在大脑中是否存在还远未清楚。(2)计算机和其他制品的组装方式也不同。虽然计算机的设计可以确保高级功能相对独立于硬件的变化,但大脑的功能可能很大程度上取决于低级别的细节,这些细节通常被认为与认知无关。有趣的是,这两种选择又被认为是两种建构方法的产物:对于计算机/制品来说是工程学,对于大脑来说进化。然而,Chirimuuta担心复杂生物系统的构建和功能(如大脑)被过度简化。
2.4 大脑即复杂:比喻、类比,理论还是事实?
Brette和Richards/Lillicrap(见下文)探讨的核心问题是一个语义问题,他们问道「计算机是什么?」Brette指出,在通俗和技术用语中,「计算机」被认为是一种「可编程机器」。然后,他指出,在计算机科学中没有计算机的正式定义,但他让我们关注「程序」这个概念,它被定义为「一组明确的指令,事先完全指定了系统的行为(『pro-』表示之前;『-gram』表示写入)」。
此外,Brette对算法和大脑计算的概念的讨论十分得当。从进化和哲学的角度来看,有两个深刻的问题与该概念有关:「大脑程序(brain program)是什么?」和「谁来『编程』大脑?」。这些问题的推理似乎导致了一个逻辑结果:「……大脑可能不是计算机,因为它不是真正可编程的。」 Brette提供了隐喻和类比的定义,并得出结论,认为大脑-计算机的隐喻似乎没什么用处,甚至是误导性的,因为它提供了一种简化认知和行为的观点。这个结论与Richards和Lillicrap的观点形成了鲜明的对比。
下一篇论文涉及与大脑-计算机比较的其他一些问题。这些问题可能涉及语义误解(该领域的正式定义)或有关计算机或大脑所能做的导致误解的假设。Bongard和Levin以及Richards和Lillicrap的论文涉及影响所有定义的根本问题。概念的定义很大程度上带有产生它们的领域(例如计算机科学、工程学或神经生物学)的烙印。这种束缚影响了我们构思计算机或大脑所能做的可能性的方式。这里,大脑、计算机和机器是被详细分析的三个例子。接下来的三篇论文,由Fraser等人、Roli等人和Danchin/Fenton撰写,明确阐述了软件(算法运行)以及大脑和计算机如何处理信息。所有作者都讨论了人工智能能够在多大程度上复制生物体的行为。尽管对人工智能的潜力存在普遍的乐观态度,这些作者引入了一些注意事项;这些注意事项不由得使人们质疑机器「模仿」人类行为的可能性。「智能体(Agency)」和「自指」成为明显的绊脚石。本节的最后一篇论文由Davis撰写,提出了由这种类比所引发的一系列问题,并询问自己(以及该领域的研究者)这些问题在多大程度上得到了回答,以及答案将对辩论产生何种影响。
2.5 生物不是(20世纪的)机器:从现代机器行为科学的角度更新机械隐喻
长期以来,人们一直认为生命和机器是根本不同的实体,前者不能归结(reduce)为后者(Nicholson,2013)。Bongard和Levin 认为,这种二分法大多基于一种旧的机器概念,即17到19世纪的观念,这种观念没有考虑到人工智能、生物工程等学科的现代发展。在这种背景下,作者重新审视这个问题,并问:「一个合适的机器隐喻是否足够适用于生物学以促进实验和概念进展?」理解这个问题的路径,要从明确定义机器以及它们的特性开始,再到批判性地评估现代科技告诉我们的这些特性。这些特性是否明确区分了活的(alive,或进化的evolved)和工程的(engineered)「事物」之间的差异?作者提到,上述学科的现代发展使得这两个「系统」之间的清晰分隔越来越难以维持,且随着现代工程学的进展,边界变得更加模糊。作者强调,对生命体所关联的特性进行分析后新开发的机器清楚地表明,曾经被认为是明显不同的系统之间的界限现在变得模糊了。在最后,他们试图对机器(machine)提供一个新的工作定义(working definition,意为发展中的定义),以容纳我们获得的所有新见解。
2.6 脑机隐喻之争毫无意义:语义问题
Richards和Lillicrap强调了一个事实,即不同的学科对「计算机」的定义不同,具体而言,他们将计算机科学中使用的定义与计算机科学之外的学科中使用的定义进行了对比。根据这个论点,关于大脑-计算机隐喻的大部分争论只是语义上的分歧。那么这样就结束了争论吗?
尽管「计算机」的通俗用法很简单——「人造设备(笔记本电脑,智能手机等)对输入进行顺序处理以产生输出」,但计算机科学中的概念却并非如此。作者对计算机科学中「计算机」的概念进行了深入分析,这个定义基于其他两个概念,「算法」和「可计算函数」。「算法」可以非正式地定义为一系列有限的逻辑步骤,机械地解决问题。「可计算函数」是「任何值可以使用算法确定的函数」。算法的正式定义是分别由数学家Alan Turing和Alonzo Church于1936-1937年独立发展的,作者介绍了Church-Turing论题。
作者提供了「计算机」的正式定义:一种「物理机器,可以实现算法以解决可计算函数」(a “physical machinery that can implement algorithms in order to solve computable functions.”)。他们强调这个定义「非常重要,因为它是计算神经科学和人工智能研究的基础」。作者还描述了这个定义在大脑方面的应用,并讨论了其局限性。
总之,Richards 和 Lillicrap 希望我们对比计算机科学内外两个定义。如果采用计算机科学的定义,「人们可以……简单地问,大脑是什么类型的计算机?」然而,「……如果采用计算机科学之外的定义,那么大脑就不是计算机,而且可以说,计算机是大脑的一种非常糟糕的隐喻。」
那么,辩论结束了吗?
2.7 为什么大脑(而不是计算机)能理解说谎者悖论?
人类大脑的一个特性是能够意识到自身的存在。这似乎是与其他标准机器(包括计算机)的一个显而易见的不同。Fraser和他的合作者(Fraser,本期)通过自指陈述(self-reference statements)为此提出了有力的论据,这些陈述无法被计算机解决。他们明确地陈述了这个案例:像「现在说的这句话是假的」这样的口头陈述被我们的大脑「理解」了。然而,计算机面对这种陈述时,会陷入一个「无限循环」。大脑是如何处理上述悖论的呢?Fraser等人提出了一个优雅的动力学模型(dynamical model),其中大脑由相互作用的单元(模块)组成,它们沿着时间轴运动(奇异循环模型 strange loop model,指在层级系统中单向移动时能回到起点)。该模型提出了大脑处理/解决悖论的一种方法,即通过将不一致性的分析延伸到时间上(沿着这个轴解卷deconvolute)。大脑能够通过时域化(temporalize)应对这个悖论。这避免了系统(大脑)陷入无限循环中。

2.8 生物如何认识世界:通用人工智能的基本局限性
Roli、Jaeger和Kauffman的焦点放在了人工智能(AI)和人工生命(Artificial Life,Alife)领域。自然智能和通用人工智能(AGI)的概念被对比,后者被定义为「把『分析、创造和实践智能』相结合的能力」。AI和Alife的最终目标是创建一个计算机或机械系统(AI-ALife-agent;例如,机器人),能够自主地(即无需人类干预)识别、评估和利用新的替代机会(称为可供性 affordances),以便以类似于自然生物(自主生物体)的方式进化和创新。这里定义的可供性是指「达成目标的机会或障碍」。作者认为,当前的AI算法不允许AI和Alife智能体「以与自然进化等同的方式进化和创新」,因为它们无法超越其预定义的可能性空间(space of possibilities,由人类设计者确定)。此外,他们还表明,在生物学和AI研究中,「智能体」这个术语指的是根本不同的概念。
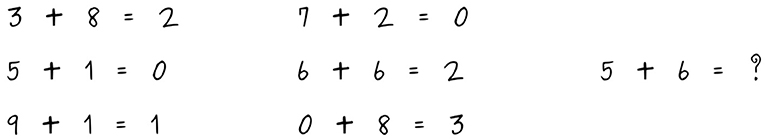
图4. 数学系咖啡室里留下的一篇论文中发现的谜语。
作为对他们立场的可能反对意见,作者提到了(i)深度学习算法和(ii)人工智能系统的不可预测性(例如,下棋、创作音乐);他们对这两个问题进行了回应并予以驳斥。
最后,作者引用了William Byers和Roger Penrose的工作,使用数学创造力的概念区分了自然智能和人工智能的能力,这种创造力「不是来自算法思维(algorithmic thought )而是通过洞察力(insight)产生的」,这种区分不是正式的,涉及到框架的转换。
2.9 从模拟到数字计算:智人的大脑是不是一台图灵机?
Danchin和Fenton的论文将神经生物学和计算机科学的概念进行了相关性分析。论文还绘制了计算、基因组学和物种进化之间的相似之处。该论文的重点(在标题中明确说明)是围绕图灵机(TM)的概念建立的;论文的一部分专门讨论了这一概念。值得注意的是,图灵机概念也被转化(transpose)到了生物学领域中。
为了探索大脑和计算机之间的潜在类比,首先需要解决一个语义问题:计算的定义。论文对比了模拟计算和数字计算的差异,强调模拟计算实现了各种反馈(feedback)和前馈(feedforward)回路,而数字计算则利用了递归过程(recursive process)。递归是这篇论文的一个核心概念,它是图灵机数字世界的一个特征。递归允许一个过程的一步(例如,图灵机的一组规则)调用自身。「机械装置通常是确定性的和可预测的,而涉及递归的计算是确定性的,但不一定是可预测的。」大脑确实进行某种形式的计算,「具有显著的效率,但这种计算基于细胞组成的网络结构,这些细胞组织成相互叠加的层,这赋予表面积/体积比例的重要性……这种计算属于模拟计算家族」(A. Danchin)。
在一张非常实用的表格中,作者比较了图灵机、数字计算机和人脑的关键特征。作者得出结论,大脑不是数字计算机。然而,他们推测,在人类的进化时间跨度上,最近的语言的发明(尤其是写作的发明,大约在6000年前)可能构成了将「人脑进化为真正(但缓慢)的图灵机」的一步。
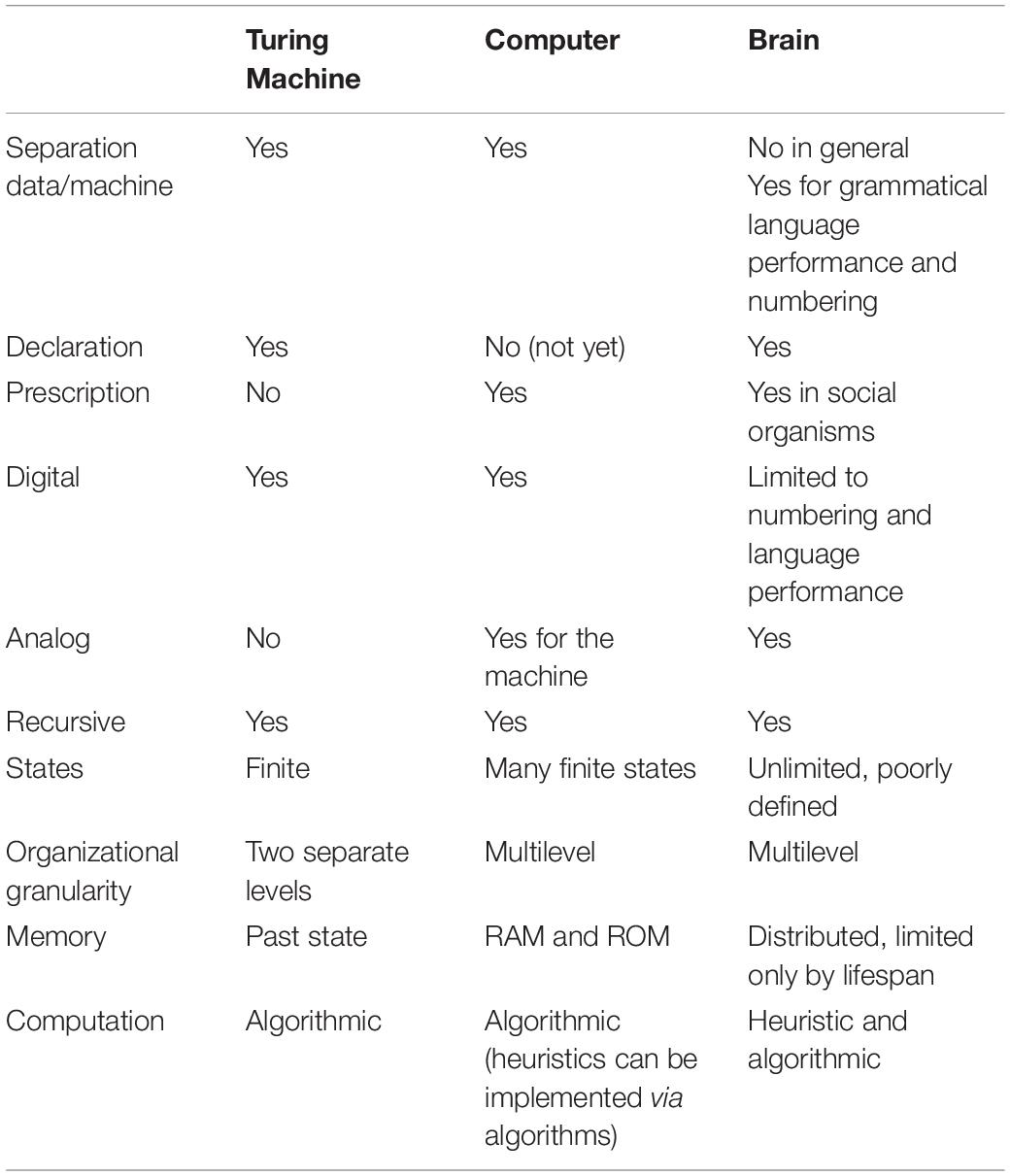
表2. 图灵机、数字计算机和人脑对比
2.10 脑即计算机的隐喻
戴维斯的简短论文提出了一系列与脑机类比相关的问题。他的重点在于大脑过程的编程。大脑用什么样的算法在世界中导航?计算机能否模拟这些算法?戴维斯建议,现代优化算法(网络训练)应该提供了一条途径,可以改进较早的数值计算。他最后提出了一个引人注目的假设:意识可能起到「大脑操作系统的接口」的作用。这绝对是计算机和人工智能领域尚未解决的问题。
Carlos Gershenson在这篇论文中引入了一种不同的视角,通过将智能的概念,特别是群体(集群)智能与信息处理理论联系起来。
2.11 作为信息处理的智能:大脑、群体和计算机
这篇论文的主要观点在标题中已经表明:它关注以信息处理为基础的智能研究。这种方法可以应用于大脑(单个和集体)和机器。
首要的问题是:人们对智能没有达成共识的定义,语义问题再次浮现。Gershenson将智能的定义建立在信息处理的基础之上:「如果一个智能体能使用信息提高其『满意度』,则可以将其描述为智能…目标包括可持续性、生存、幸福、权力、控制和理解。」 在之前的工作中,Gershenson建议使用信息的度量(measures of information)作为研究复杂性、涌现(emergence)、自组织(self-organization)、稳态(homeostasis)和自生系统(autopoiesis)的工具(Fernandez,2014);在这里,他旨在将这种方法扩展到认知系统,包括大脑和计算机。
信息是一个新的语义挑战。Gershenson在引用Shannon的经典作品时提出了信息的定义。我们的注意力集中在正在传输的信息的意义上;在这种情况下,拉普拉斯妖(Laplace daemon)的失败(译者注:即「决定论」的失败),有助于确定生物学及其他学科中一个关键的、被忽视的概念:各种尺度和参照系的存在应改变用于解释某一特定现象的模型和假设。「即使完全知道系统组成部分的状态,预测也是有限的,因为相互作用会产生新的信息。」
本文中提出了有关「单一大脑」的智能和的集群的集体智能之间的刺激性比较。以昆虫群体为例,它们可以被描述为信息处理系统,其处理是分布式的。Gershenson比较了大脑和集群的认知架构,并确定了两者之间的关键特征:「大脑信息处理的速度和可扩展性比群体高得多:神经元可以在毫秒的时间尺度内相互作用,而昆虫在秒的时间尺度内相互作用,实际上这极大地限制了集群在信息处理方面的能力。」
最后,「智能作为信息处理」的思路被用作一种隐喻,以理解其演化和生态。关于生态(选择性)压力对智能的演化和生态系统复杂性的影响,作者的可能会有不同的观点。
总之,虽然「将大脑比作计算机的隐喻不适用于研究普遍的集体智能(collective intelligence),更不用说集群智能(swarm intelligence)了……」,但是因为「计算可以被理解为信息的转化(Gershenson,2012),广义上理解为处理信息的机器可以是一个有用的隐喻……」
Graham在比较大脑和计算机时引入了一个更复杂的元素。他指出,我们不应该将大脑与一台计算机进行比较;实际上,后者的运行更好地由相互连接的计算机集合(即互联网)来代表。他指出了一个相关问题,即那些支持「强」大脑-计算机类比的人无法解决的信息路由问题(即信息在大脑和计算机内的流动方式,以及它如何从输入指向输出)。
2.12 互联网工程的九点启示——帮助我们理解脑网络通信
Daniel Graham分析了计算机-大脑隐喻的适用性(另见Graham,2021);它被实例化为他所称的神经元组件的「表征(representational)」视图。根据Graham的说法,这个隐喻是有用的,但也是不完整的。虽然他同意大脑进行了一些「计算」,但他认为大脑本身可以被看作是表征和通信活动的结果。强调大脑中纯数学运算的重要性以及它们在神经元电气脉冲模式中的转换,并不能完全展示大脑在执行任务时内部发生的情况。不支持严格的计算机-大脑功能(表征)类比的原因之一是它没有处理大脑神经元网络中信息流动的关键问题。信息路由和电路的重塑超越了计算机隐喻的限制。Graham建议互联网是我们(或任何人)大脑架构和功能属性的更好形象。互联网是通过清晰的路由协议构建的,具有高效的信息分布(称为小网络配置 small-network configuration,Sporns和Honey,2006;但请参见Hilgetag和Goulas,2016年的批判性观点),以及它们连续的连通性重塑(以及生长)。这些属性应该让我们想起大脑如何被构建以及大脑如何路由信息。整合信号处理和路由策略的观点应该给我们一个更细致的大脑活动视图。
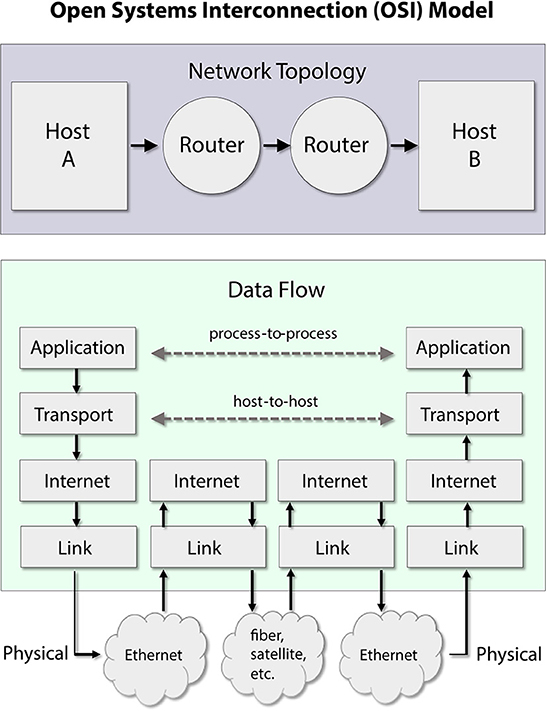
图5. 基于开放系统互连(OSI)模型的互联网协议栈的简化概念设计。
3. 讨论延伸
3. 讨论延伸
3.1 脑机类比:「将信将疑」
就像科学假说一样,隐喻和类比是短暂的,总是会随着技术进步而调整。大脑-计算机通常被称为隐喻,但实际上应该被视为类比。我们在这里指出,隐喻和类比是两个不同的概念,不能被用作同义词(见上文)。虽然我们认为隐喻的一个显著特征是「视觉洞见(visual insight)」(具有生动的视觉形象),但类比的概念主要与「功能」相关。简而言之,隐喻没有解释能力,而类比则有,因为在对一个系统的功能知识被迁移到类似的系统时,可以导致理解和发现。
脑机类比在科学界引发了激烈的争论;一些人从字面意义上理解,而类比的含义只暗示了部分属性的重叠。事实上,类比或隐喻很可能是不可避免的(并且经常用作认知工具;参见Lakoff和Johnson,1980;Gomez-Marin,2022)。隐喻根植于我们知道和/或操作的事物。在这个意义上,理解许多事物的唯一方法是用我们理解的术语描述现象。在从「物理现象」到「理解它」的过程中,隐喻/类比总是出现。我们假设只有那些由感官领悟的事物才能被认为是「原始的(original)」(总是假定我们的感官没有欺骗我们)。隐喻可能是我们「理解」的唯一东西,这是因为它们根植于我们的感性体验。Kuhn本人似乎承认隐喻的重要性,他声称:「隐喻对于连接科学语言和世界非常重要,但是这些连接并不是永久性的。随着理论的变化,相关的隐喻和连接科学术语和自然的相似性网络的部分也在变化」(1993)。
在这种情况下,问隐喻和类比是否真正有用或具误导性是错误的问题。Yohan (2012)恰当地指出:「没有人能声称知道隐喻是如何起作用的……我们如何形成它们,以及我们如何决定它们是否成功。」 同样的道理,我们确信隐喻和类比是否「正确或错误」对它们在科学中的作用是完全无关紧要的。这种态度的一个绝妙例子来自于Niels Bohr的马蹄铁轶事(horseshoe anecdote):一个朋友问他是否相信家门上挂着的马蹄铁会带来好运。Bohr回答说「绝对不!」,朋友回答:「但是他们说,即使你不相信它,它也能起作用。」
有用性似乎是隐喻更合适的形容词;因为它们为所分析的现象提供了线索。
此外,从数学的角度来看,威廉·拜尔斯(William Byers)认为,「许多重要的数学思想本质上是隐喻性的」,并强调「隐喻和思想之间的密切关系。隐喻就像思想一样,源于创造性的行为」(Byers,2010,第240页)。此外,Byers指出:「通常情况下,大多数笼统的猜想(sweeping conjectures)在被研究期间就被证明是「错误」的,因为它们需要修改。尽管如此,它们仍然可能非常有价值。整个数学研究通常都是这样进行的——充满灵感的错误之路……即使是「错误」的思想仍然可以有价值。」
本专题的一些文章涉及到在神经科学和计算机科学之间的交叉领域中对隐喻的使用。在这个背景下,需要再次强调,隐喻在本质上是必不可少的,但是隐喻也是暂时的,因为新的、更新的数据可能会导致对其他隐喻的阐述(或先前版本的迭代版本)更加适合当前情况。此外,在没有新数据积累的情况下,社会学或认识论的变化在某些时候也可能是新隐喻的来源。此外,Gershenson 提醒我们,在学科内部,不同的隐喻可以用于不同的目的和不同的环境中。例如,利用唯一的隐喻(如下文所述)可能不是解释某些复杂结构(比如大脑)最有效的途径。
在前面的段落中,我们提出了两个特征,可用于表征和区分隐喻和类比。在我们看来,隐喻是从一种视觉形象发展而来的,这种形象提供科学洞察力的创造性力量。再次引用数学方面的观点,Ivar Ekeland强调了数学思想与「某些图像」的关系以及这些图像的力量:「在科学的历史发展中,某些视觉表达方式可以成为产生新的和成功的想法的有用工具。在早期阶段,当它传达的思想仍然是创新和成功的,它有推动进步的力量;而当推动力消失,旧理论的重复阻止新思想出现时,它就变成了一种阻碍的力量」(Ekeland,1988,第9页)。
正如丹尼斯·诺布尔所说:「不同、甚至相互竞争的隐喻可以阐明同一情况的不同方面,尽管这些隐喻本身可能不兼容,但每个隐喻都可能是正确的。……隐喻之间在启发性、简单性、美感、创造力等标准上展开竞争。隐喻的创造是一门艺术而不是一门科学,就像其他艺术形式一样,艺术家不一定是最好的解释者」(Noble,2006)。我们完全赞同这些观点。
至于类比,我们认为「功能」这一标准可能被视为其最具特征的属性,这是一种具有解释力的属性。例如,在这个专题中,Daniel Graham提出了互联网作为大脑的一个新隐喻。根据我们的定义(见上文),在这种情况下,互联网既起到了隐喻的作用,又起到了类比的作用。
在这一节的结尾,我们需要谨慎地指出,由于与学习或寻求解释相关的心理因素,使用隐喻可能是不可避免和必要的(Hoffman,1980)。不使用隐喻的替代方案将导致对事实的简单描述。在心灵哲学(或我们在这里的讨论)中,这意味着对大脑中生理状态的「纯」描述。无论这个描述是否有任何「信息内容(information content)」,本综述的作者认为,它非常少,甚至没有。我们无法设想在科学中使用隐喻的任何有效的替代方法。
3.2 大脑的理论
我们想强调的一种观念是将大脑视为一个可以在「不同尺度」上进行分析的结构,该结构的功能可能分布在不同领域,并涉及不同组件。在这个意义上,我们认为寻找「大脑的终极理论」是错误的,因为更好的描述应该包括关于如何建立和整合不同层次的结构组织(包括各种功能域 functional domains)的解释。正如下文所述,探索「大脑的不同理论」可能更为合适,这也是我们下面提出的探索性努力的更合适的名称。
在这种情况下,我们想通过系统方法,用一种不同的视角分析脑机类比。其中,每一种类比对应着不同的组织水平。我们认为,一种试图类比组件(components,或模块 modules,见下文)的理论应比涵盖复杂结构(如大脑)的单一理论更具生产力。这里的基本假设是,大脑是组件/模块的(非线性)总和,这些组件/模块并置(juxtaposed)以执行或推进某些心理任务。这不是毫无根据的假设,因为当前的神经科学数据已经证明了大脑中许多结构的模块化,它们都是进化的历史产物。从神经元亚型的共性到特定神经元回路的保守性或皮层区域的分布,许多动物的大脑都共享中都存在负责特定功能的结构,这种特性已被认为是跨类同源(homologous across taxa)的(Schlosser, 2018; Barsotti et al.,2021; Tosches, 2021)。
实际上,大脑和其他器官或组织一样,在不同层次上有不同的组织结构,一个层次上的模块对下一层做出贡献;这暗示着一个平行的讨论,即类比大脑结构和更经典的跨尺度生物同源性讨论(蛋白质、细胞、器官等)。(参看《模块化认知:演化如何自下而上涌现出智能?》)
在这个框架中,大脑的层次化结构使我们重新关注最能代表不同尺度的类比(即在一个层次上是计算机,在另一个层次上是收音机,在下面是全息照片,在上面是互联网等)。这并不意味着将整个大脑的问题搁置一边,而是找到更好的(有用的)类比,以代表参与其构建的那些较低级别的模块,并将模块用作已被认可的功能单元(例如神经元或神经回路)。这里需要引入一个警示性的注释:我们并不声称对大脑模块化的解释是有力/严格的,因为我们知道大脑有明显的分布式功能和可塑性,以及随时间变化的刺激表征。实际上,模块的分布式和灵活组织特性使我们能够集成各个层次。
从某种意义上说,正如Cobb所解释的那样,我们对大脑发育和其功能的假设在每个历史时期都依赖于那时对该系统的认识,如液压学和电学的发展带来了对大脑印象的同步更新。计算和算法推进了对大脑计算思想的发展,尽管后来的观点认为复杂的计算是在单个神经元的突触上进行的。类比有时从一个层次变化到另一个层次,上述的大脑计算形象就是一个例子。同理,网络类比从少数神经元的局部连接(反射弧)转变到涉及特定任务的大脑区域,再到整个大脑或其群体。
总之,我们建议重新评估我们使用类比的方式,以便更好地了解每个层次是如何组织的,以及在特定层次集成不同模块如何产生更高层次的(涌现)特性。此外,我们鼓励引入不同的类比,以最好地代表不同层次的建构(这也许能解释复杂系统中的每个单一组件,避免总体类比的陷阱)。我们应该注意到,在生物有机体构建的「部分(parts)」可以归因于偶然性(漂移)、物理化学规律(自组织)、涌现现象或适应性过程。所有这些构建原则与我们对有机体的纯结构观点没有任何关系。在这里,我们的建议基于对结构本身的多层次分析,但不涉及它们的发展性组装。也许可以在未来考虑这种最后的近似,它以类比搜索的精炼形式出现。
3.3 科学创造力
在本期特刊中,我们为什么对创造力(creativity)感兴趣?因为如果我们问「计算机能思考吗?」,接下来的问题就应该是「计算机能创造吗?」,而创造行为(不论是在科学还是艺术方面)源自对美学元素(esthetic element)的认识。对创造力及其源头的思考有着悠久的历史,虽然哲学家们对创造力的定义和内容仍未达成共识(参见 Erden, 2010),但一些原则(tenet)被公认为与创造性行为有关(如自由、潜力、独创性等)。此外,一些哲学家还认为,创造,实际上是一个有一些特定要求的过程(McGilchrist,2021)。这些要求包括(i)创造力(允许想法产生;识别模式等)(ii)宽容(作为想法进一步发展的条件)以及(iii)转化倾向(将洞察力延续一段时间,直到最终的「创造性行为」)。这些并不是其他(不一定具有创造性的)过程的组成部分(例如解决问题)。从后一种意义上说,“创作过程背后没有算法”已经得到了证明。许多作者都认为,隐喻可以扩展创造性思维,也就是说,对隐喻的分析成为理解科学创造力的关键组成部分。那么,我们如何进行分析?
对隐喻和类比的分析,其基础在于理解它们的来源。创造力显然是一个来源。但是,「真正的创造力」在科学中的来源是什么呢?毋庸置疑,科学史告诉我们,机遇(chance)在科学创造力中起着重要作用,并将继续发挥作用。除了传统观点,近年来还探索了更具创新性的路径。这是对大脑-计算机隐喻讨论的一个重要议题。
但首先我们需要定义什么是创造力,以及创造力和隐喻之间的关系是什么?
在形式(formal)科学和自然科学中,关于创造力的问题已经被广泛地讨论,它们主要集中在数学和物理学领域。数学家William Byers区分了数学中的两种思维类型:算法思维(基于逻辑运算)是平凡(trivial)的,而深刻的思维则是具有创造性的(Byers,2010)。
Byers提出了一些深刻的问题:「计算机能否被编程来区分琐碎的和深刻的事物?」……「计算机能做数学吗?」……「数学是算法吗?」 因此,他不可避免地面临「问题(THE question)」,这个问题最初是图灵在1950年提出的:「计算机能思考吗?」他说「……相当于问题是:[人类]的思想能用算法表示吗?」 如果按照Byers的说法,「人类的创造力涉及到思想(idea)、歧义(ambiguity)、悖论(paradox)、深度和复杂性」,那么创造力的行为(这是一个非常罕见的事件)可以类比于生物进化事件,因为(正如一些作者所指出的那样)不可能预测思想将来如何发展。思想似乎随着时间的推移、通过文化(culture)进一步发展(Gabora & Kaufman, 2010)。
我们在这里提议考虑创造力的另一个方面,我们认为这个方面非常相关,但仍然没有得到充分的认可:它是研究人员在面对现象时可能会感受到的惊奇的感觉(amazement,法语中称为émerveillement)。这种感觉可能是唤醒科学家并启发他/她思考的必要和充分条件。此外,我们还想提出,在可能的「émerveillement」的促进下,科学创造力可能取决于来自不同学科的观点的整合(许多科学家在被问及自己的工作时已经提到了这一点)。在这种情况下,我们推测,从一个领域向另一个领域「转移隐喻(transferring metaphors)」可能是新思想的优秀来源,也有助于创造性的行为。在一个领域作为理解工具的隐喻应该能够对另一个领域的其他现实方面有所启示。这种思维上的「转移」可以是有成果的,并因此导致对不相关现象的「理解」。
在脑机类比的背景下,人类创造力与「人工智能创造力」的比较自然而然地出现。争论人工智能是否在人类创造力中发挥作用,无疑是时代所需。
很显然,人类创造力受文化传统(语境)或不同思想之间的联系(即来自不同/不同领域的思想)的影响。这可能表明,人工智能在模仿人类创造力的过程上存在一些固有的困难。
按照上述说法,我们可以说,计算机不具备创造力,无法产生「创造性行为」,因为创造可能不是纯粹的算法过程。
正如Byers(2010)支持人类创造力的优越性的文字:「数学思维可以很简单,也可以很复杂,但大多数情况下是不平凡的。另一方面,计算机思维可能非常冗长和复杂,但它本质上仍是平凡的。」 Roli等人也支持在数学中的创造力「…并不是来自算法思维,而是通过洞察力获得的」,基于这样的论点,即由人类设计的人工智能算法对新的参考框架,无法识别、评估和利用(适应于)新的「环境」选择(称为affordances)(Roli等人,2022)。
支持AI潜力的人描述了它在自主创意过程或在人类干预下的表现,主要在写作、音乐和绘画等领域(例如,Zylinska,2020;Jukebox,https://openai.com/blog/jukebox/ )但在科学发现中也有涉及(例如,通过机器和深度学习技术进行蛋白质折叠预测;参见Gil et al.,2014和Callaway,2020)。
其他作者持更加乐观的看法,认为AI可以具备创造力(例如Boden,2003; Forbes,2020)。这些作者根据经典心理学原则(例如Boden,1992)认为创造力具有不同的来源,并将其分类为:组合性创造力(结合熟悉的想法;例如诗意的形象)、探索性创造力(在文化传统中出现;例如烹饪食谱)和变革性创造力(打破旧规则;例如立体主义)。虽然组合性和探索性创造力的组成部分可以被AI模仿,但计算中变革性创造力的可能性对AI来说似乎更为棘手。打破规则似乎是人类特有的能力,因为对这些新规则的最终接受取决于「价值判断」(只有满足某些新需求的结果才被纳入)。当然,拥有进化机制(具有选择性机制)的程序现在已经成为现实,这使得「变革(transformation)」成为可能。在这里,价值是一个重要的选择因素,但AI目前很难实现。我们应该记住,批判性思维仍然是创造过程的核心。人类能够审查他们的想法/创作,计算机则不能。总之,这些作者建议,通过确立大脑可能是一个精妙复杂的机器的观点,最终摒弃笛卡尔的心物二元论(Cartesian dualism,译者注:笛卡尔的心物二元论是一种认为心灵和物质是两种完全不同的实体,互不相干、各自独立、平行存在的本体论学说。心灵的本质是思想,根据自由意志而活动;物质的本质是广延,遵循自然规律而运动。笛卡尔提出这种观点是为了把物质限定在可以用数学模型描述的范围内,而把思想作为一种无形体,脱离身体和客观世界的存在)。
尽管存在这些观点,但每个科学家似乎都同意,在当前的人工智能发展状况下,我们的「机器」模仿人类创造力的能力现在仍然相当有限,尽管未来可能会迅速改变我们的观点。总之,当讨论与本特刊相关的隐喻时,应将创造力纳入关键概念。
在这篇综述文章的结尾,我们还要提到的是:不应忽视美(以及一般美学)在科学创造力中的作用。
创造力与美学密不可分。评价创造力总是需要美的评判。例如,根据康德的经典论述:「美学不是创造力的目标,而是其基本组成部分。」
数学家和物理学家们承认美(以及美学)在科学中的至关重要性,他们包括G·W·莱布尼兹(G. W. Leibniz)、亨利·庞加莱(H. Poincaré)、阿尔伯特·爱因斯坦(A. Einstein)、G·H·哈代(G. H. Hardy)、P·A·M·狄拉克(P. A. M. Dirac)、M·盖尔曼(M. Gell-Mann)等人。1910年,庞加莱写道:「…… 数学美感、数字和形式的和谐感、几何优雅感。这是一种真正的审美感觉,所有真正的数学家都知道,它肯定属于情感灵敏度。那么,我们认为具有美感和优雅感的数学实体是什么,它们能够在我们中引起某种美学情感?」(庞加莱,1910)。狄拉克在一句著名而常被引用的话中大胆地说:「在公式中具有美感比公式能够拟合实验更加重要」(狄拉克,1963)。在哈代的话中:「数学家的图案,就像画家或诗人的图案一样,必须是美丽的。」最后,浪漫主义英国诗人济慈(John Keats)通过两行极具影响力的诗句完美地概括了美学方法对人类知识的普遍性:「想象中的美丽必须是真理」(1817),「美是真理,真理是美。这是你们所知道的一切,也是你们需要知道的一切」(1884;引自Keats,2015)。最近,Yang and Lu(2022)探讨了计算机科学、创造力和美学之间的关系,并提出了一种框架,使用计算方法将创造力和美学联系起来。
很明显,美的概念在不同文化和个体之间存在着很大差异,这是显而易见的。实际上,我们通常认为美学质量(esthetic quality)的概念是难以捉摸的。但问题在于,正如著名画家Bruegel的画作「The blind conducts the blind」所寓意的那样,每个人都在寻找「绝对」的美的定义,而这正是错误之处。就科学而言,美的最关键的一点是,所有的美的含义和定义都是等价的,都能达到相同的目的:为科学发现指引道路。美是无法定义的,因为它在观察者的眼中,这并不重要。任何美学标准能够发挥作用的机会都是不可忽视的。同样,对于隐喻和类比也是如此。
参考文献
(参考文献可上下滑动查看)
NeuroAI读书会
详情请见:
NeuroAI 读书会启动:探索神经科学与人工智能的前沿交叉领域
后ChatGPT读书会招募中
2022年11月30日,一个现象级应用程序诞生于互联网,这就是OpenAI开发的ChatGPT。从问答到写程序,从提取摘要到论文写作,ChatGPT展现出了多样化的通用智能。于是,微软、谷歌、百度、阿里、讯飞,互联网大佬们纷纷摩拳擦掌准备入场……但是,请先冷静一下…… 现在 all in 大语言模型是否真的合适?要知道,ChatGPT的背后其实就是深度学习+大数据+大模型,而这些要素早在5年前的AlphaGo时期就已经开始火热了。5年前没有抓住机遇,现在又凭什么可以搭上大语言模型这趟列车呢?
集智俱乐部特别组织“后 ChatGPT”读书会,由北师大教授、集智俱乐部创始人张江老师联合肖达、李嫣然、崔鹏、侯月源、钟翰廷、卢燚等多位老师共同发起,旨在系统性地梳理ChatGPT技术,并发现其弱点与短板。本系列读书会线上进行,2023年3月3日开始,每周五晚 19:00-21:00,欢迎报名交流。

推荐阅读
微信扫一扫,分享到朋友圈











