演化工程师:生命与环境如何相互塑造?

导语
在演化的长河中,生命与环境持续地相互塑造。环境以自然选择的力量给生物压力,而生物也会通过调整自身所处的生态位进而改变演化压力,这被称为生态位构建理论。尽管生态位构建理论尚未成为当前主流的演化理论,但其大幅拓展了对生物适应性的理解,并且为人与环境的关系、社会经济和组织管理等问题提供了新的启发。

Kevin Laland、Lynn Chiu | 作者
赵雨亭 | 译者
张澳 | 审校
邓一雪 | 编辑
1. 自然选择:登顶还是移山?
1. 自然选择:登顶还是移山?
人类正在塑造地球生命演化的未来。我们的活动不仅造成大量物种灭绝,还迫使动植物与微生物适应我们的人造世界——例如,城市鸟类如今改变了自己的鸣叫,因为更高的音调似乎可以使歌声传播更远。在文化知识与工程技术提高人类改造环境的能力的同时,其它物种也都在改造环境。在培养皿中涂布细菌,它们将产生营养丰富的代谢产物——新的细菌菌株可以利用这些副产品,迅速使多种多样的微生物种群进行演化[1]。海狸创造的池塘为鸭子提供了繁殖地;残留的蜘蛛网被地上的昆虫开发为冬眠地;植物通过其根部分泌的物质来改造土壤。这些生物改变环境的方式反过来改变了它们和其他物种在生存与繁殖中所面临的演化压力。换句话说,它们的行为使自然选择产生偏向。所有物种都会进行这种被称为“生态位构建(niche construction)”的过程,虽然它们造成的影响远不如人类剧烈而广泛。
然而,生态位构建的理论[2]在演化生物学家中引起争议,部分原因是传统上认为自然选择是“盲目的”:人们认为,对生物体上千年的调整是为了使其适应各自的生态位,而不是为了达成生物体各自的目标。人类经历了同样的改造,但相比于为了适应预设的生态位而演化,我们更加主动,更像是在改造环境而使其适于生存。我们的大脑已经演化为适合处理语言编码的信息与知识,因为我们建立了丰富的文化领域,并随后对其适应。我们驯化了动植物,将其纳入我们的食物中,从而触发了代谢此类食物的基因选择。我们创造了农业,促使人口增长,无意中筛选了对群体疾病(如伤寒或霍乱)的抵抗物质。我们的父辈不仅将基因传递给我们,同时还留下了其改造后的世界。这种生态遗产意味着人类并非响应静态的环境而演化,而是塑造该环境以缓解或加剧其施加的某种选择压力。演化生物学家理查德·莱文斯(Richard Levins)和理查德·列旺汀(Richard Lewontin)指出[3],“有机体既是自然选择的对象,又是选择条件的创造者,从而影响着自身的演化。”
但这种观点仍有争议:专家们对其它生物通过改变环境来指导演化的程度产生了分歧。地形的比喻可以帮助理解演化过程。我们可以将演化视为适应性景观(adaptive landscapes,也译为适应性地形图),该图揭示了生物体的适应性(其生存和繁殖能力)与一种或多种生物体性状之间的关系。例如,在一张鸟群适应性景观中,一个轴可能代表体型大小,另一个轴可能代表尾巴长度(或者可以用相关基因的频率来替代)。该地形图为三维空间中的曲面,具有更高适应性的鸟——即得分最高的那些——具有更优良的体型与尾巴长度组合,而适应性较差的鸟则处于曲线低谷。曲面最高峰的区域代表具有最佳体型和尾巴长度的鸟,同时也是生存能力最强的鸟。
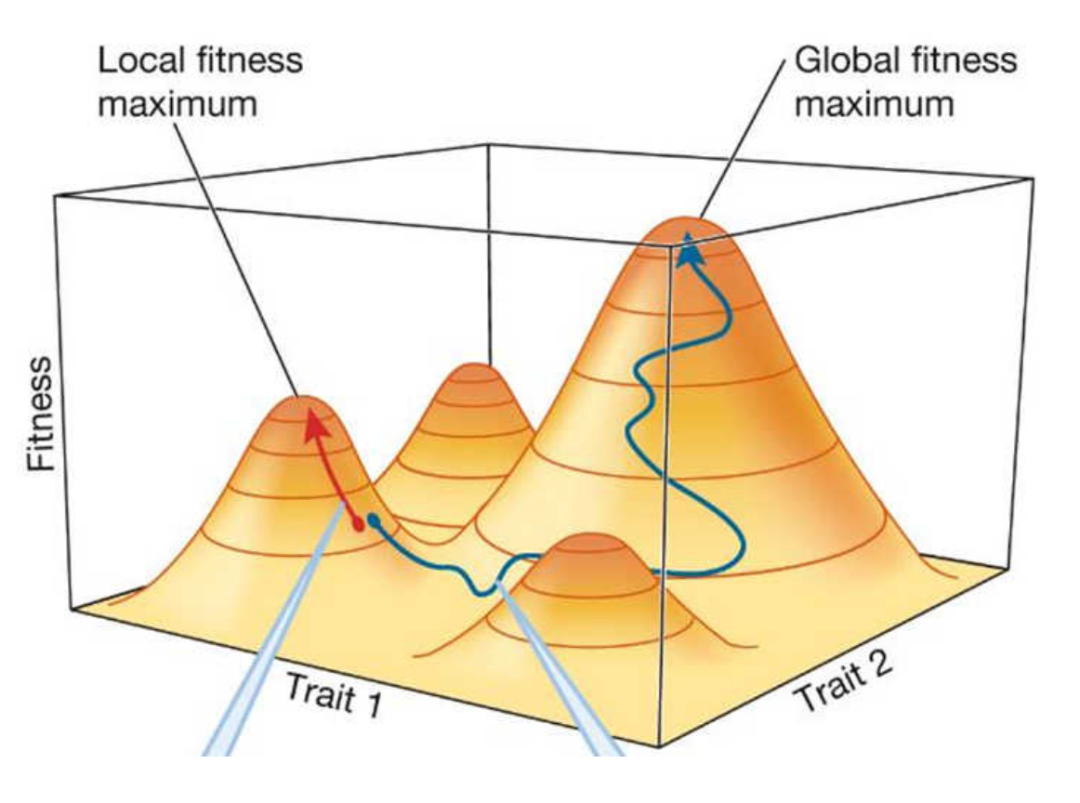
图1. 三维的适应性景观
从演化尺度上看,鸟类种群具有一系列特征,每种组合在地形图中对应不同的点——通过自然选择,种群将逐渐收敛到最适合当地栖息地的特征,通过种群演化而在适应性景观上攀升至适应性的局部高峰。然而登顶之路不止一条:可以调整自己的性状以到达顶峰,也可以移动山峰以适应自身的需求。
2. 蚯蚓的进击:从适应到入侵
2. 蚯蚓的进击:从适应到入侵
小巧而有力的生态位构造者随处可见。低调的蚯蚓就是一位“生态系统工程师[4]”,它是一种会改变物理与化学环境的物种,并常以改变生态系统功能的方式改造环境。奇怪的是,尽管在生态学上蚯蚓是成功的,但在解剖学上,它们实际并不适合在陆地上生存。尽管已经在陆地上生活了数千万年(甚至可能数亿年),但它们仍保留着从淡水中演化而来的基本生理特性。与大多数陆生动物相异,蚯蚓会如淡水动物般代谢产生大量尿液,这使它们极易脱水而亡——如果在晴天挖出了蚯蚓,它们通常会在短时间内干枯。那么,这样一种生理适应性如此差的动物是如何在陆地上生存这么长时间的呢?
答案是生态位构建——相比于调整生理特征以适应陆生环境,它们选择了在陆地上制造水生世界。它们的成功直接源于对自然所干的园艺活。蚯蚓通过松土减少土壤的“基质潜力”——即土壤保持水的能力。这使蚯蚓更容易通过周围环境摄取水分,以补充由于它们的排泄而造成的水分损失。换句话说,蚯蚓有效地建立了自己的储水池[5],使自己保持湿润。
就这样,蚯蚓发展了许多将环境改造为适应其自身发展的方式,并造成了与人类入侵自然类似的影响。如今欧洲蚯蚓被认为是北美森林植物群落发生变化的主要推动力[6]——并且与人们认为蚯蚓对土壤有益的固有印象相反,蚯蚓对阔叶林具有毁灭性的影响。北美本土蚯蚓种群在第四纪冰期期间几乎灭绝,在16世纪和17世纪欧洲殖民者将蚯蚓引入前,多数生态系统没有蚯蚓的存在。随后,欧洲蚯蚓迅速扩散,它们入侵落叶林并快速改变了土壤性质,使本地动植物物种生存困难。
3. 入侵导致的级联反应
3. 入侵导致的级联反应
蚯蚓将有机物作为食物或用作挖洞的材料拖入土壤中,导致枯枝落叶层迅速流失,并导致土壤表面升温、脱水。蚯蚓的活动改变了碳和氮等化学元素在整个生态系统中循环的速率——例如,加快了植物有机体的分解速度,同时它们的挖掘行为提升了土壤的通气和排水效果。这些变化通常在入侵的前十年内发生,进而引发了持续数十年的演替状态。历史上,在那些没有蚯蚓的森林中,大量碳元素被锁于森林表面的枯枝落叶层下。而当蚯蚓消耗这些枯枝落叶时,大量储存的碳元素以二氧化碳的形式被释放到大气中,就像人类使用化石燃料一样,从而导致全球变暖[7]。

遭遇蚯蚓入侵的栖息地产生了方方面面的变化,对其它物种产生了深远的影响。最近的研究发现[8],随着引入的蚯蚓种群丰富性增加,植物多样性下降,其中本土植物受到的冲击最大;随着蚯蚓生物量的增加,土壤无脊椎动物减少[9]。本土动植物遭受冲击的原因是因为它们以往适应的是不存在蚯蚓的环境。同时,某些物种受益:就像人类的侵扰一样,蚯蚓的工程促进了快速生长的细菌、草本植物与一些大型动物(例如鹿)的入侵和繁殖。蚯蚓的洞穴被其它无脊椎动物——甚至某些脊椎动物(如蝾螈)——用作巢穴[10],而这些生物的粪便为微生物活动的提供了充足条件。与蚯蚓同时出现的往往还有侵入性灌木,如沙棘和金银花。部分原因是蚯蚓通过选择性地(例如根据种子大小等性状)食用种子,改变了种子库(种子在土壤中的自然储存)的组成。从蚯蚓口中存活下来的种子随后受益于蚯蚓粪便提供的营养及物理保护,获益于此的依然是非本土植物。
蚯蚓的入侵无疑会触发森林生态系统的级联反应。一些研究人员甚至担心蚯蚓可能引发“入侵崩溃(invasion meltdown)[11]”,其中非本土物种加速促进彼此的入侵[12]。尽管不够直观,但蚯蚓自身会从其工程设计及其触发的入侵中受益,从而使它们能够在适应性景观上创造新的山峰。蚯蚓促进侵入性灌木的生长,侵入性灌木反过来促进蚯蚓种群的生长[13]。灌木产生更高质的枯枝落叶层(即较低的碳氮比)以供欧洲蚯蚓享用,使自持续的反馈闭环得以形成。蚯蚓积极参与耕作——准备土壤,改变种子库,使得其偏爱的作物肆意生长。它们与鹿、猪和微生物建立了类似的互惠关系,后者既从蚯蚓的活动中受益,也促进了蚯蚓的活动。有趣的是,蚯蚓物种的多样性与其生物量相关[14],这表明某一种蚯蚓的工程对其它各种蚯蚓均有益。
4. 生态遗产的累积
4. 生态遗产的累积
蚯蚓也会通过建造洞穴来积累演化上上的优势。目前地球上有6000多种蚯蚓,它们在生态和行为上有很大不同。然而,大多数蚯蚓并不是闲逛的“游民”,在土地上四处游荡以寻找食物, 而将遗弃的土地扔在身后。相反,蚯蚓可以建造半永久的洞穴,其存留时间通常远超蚯蚓的一生。查尔斯·达尔文曾指出,蚯蚓的洞穴并不是简单的挖掘工作,而是坚固的垂直隧道,其深度有时超过一米,他将其描述为“内衬水泥”。蚯蚓的“水泥”通过压实其分泌物形成[15],并创造了厚度足有一厘米的独特洞穴,称为“蚓触圈(drilosphere)”。每一个蚯蚓的洞穴(它们的生活区)会在土壤表面形成粪堆,既是厨房又是厕所,由枯枝落叶及其排泄物组成。蚯蚓甚至会挖掘用于产卵的侧洞。许多蚯蚓在夜间寻找食物和配偶,它们通常把尾巴末端置于洞内,以便快速响应危机。

图4. 蚯蚓错综复杂的洞穴
挖洞是一项耗费精力的活动——认领和翻修前任主人的房子要比从头开始建造自己的房子容易得多。人们通常认为蚯蚓挖的洞穴比它们自身宽度窄,但常能在大洞穴里发现小蚯蚓,这表明要么是蚯蚓幼虫继承了父母的洞穴,要么是分散的个体倾向于在空的洞穴里安家。长期的野外观察表明,洞穴被使用的时间远超蚯蚓一生的生命。因此,蚯蚓在被使用过的土壤与未被开垦的土壤中更倾向于前者也不足为奇了[16]。
改良的土壤、现成的房屋和更充分的食物供应,并不是蚯蚓留给后代的全部。每一个蚯蚓茧都继承了一个来自其父母肠道的微生物群落。蚯蚓的环境不仅仅是土壤和周围的生物,还有寄居在其消化系统和周围蚓触圈中的细菌、真菌和单细胞真核生物。这种安排同样是互利的:蚯蚓为土壤微生物提供了一个食物丰富和可移动的环境,让它们有机会在缺氧的穴中发酵并在土壤中繁殖,而这些共生体的存在增加了蚯蚓的适应性[17]。
就像人类一样,蚯蚓已经建立了自己的生态位。它们的生活环境并不是一直保持贫瘠或富庶,而是根据其内栖息者的活动呈现动态的变化。除了改变土壤,蚯蚓还如人类一般种植农作物、释放温室气体、形成聚落并向外拓展。
5. 生态位构建:刻意的设计
5. 生态位构建:刻意的设计
蚯蚓并不是通过生态位构建改善环境的唯一物种:海藻分泌的粘性化学物质会束缚沙土并稳定环境[18];丛林和松树通过散布易燃的针叶、果皮和油类物质来提升森林火灾规模[19],从而消除竞争;蚂蚁则通过喷洒酸性除草剂或种植真菌作物[20],消除不适合其居住生活的树木[21]。
动物根据自己的需求改善环境的方式是否激励了我们以新的方式思考演化的适应性景观?从最根本的生态位构建的角度来看[22],答案是“YES”:生物不单单适应环境,生物与环境两者共同指导演化。例如,海狸的影响要比它们创造的池塘要广泛得多。海狸既是消防员[23],同时也是灌溉管理人员[24]——它们极大地改变了所在的生态系统,使其能够抵御诸如火灾或是干旱等恶劣气候的干扰。这样,海狸便会改变其本地环境,从而改变选择对它们和其他物种的作用方式。海狸是反馈与因果关系循环网络的一部分,在该网络中,初期占据优势的生物体会驱动环境变化,而经优势物种调整的环境随后会选择生物体的演化方向。
生态位构建与生态和演化中的其他几个概念重叠,特别是“生态系统工程”和“生态演化动力学”。但是,生态位构建理论的独特之处在于,生物对环境的改造(生态位构建)及其对后世的遗产(生态继承)是演化过程,正是它们引发了演化的改变。
尽管大多数教科书都侧重于描述直接改变基因频率的现象——例如自然选择、突变或随机遗传漂变(偶然发生的群体遗传组成变化),演化过程的起因目前仍有争议。回到适应性景观的比喻,这些力量推动着生物群在环境中的适应性上下移动。相反,生态位构建理论则提倡更广泛的演化因果关系概念。
引用演化生物学家道格拉斯·富图玛(Douglas Futuyma)的话说[25],“传统主义者”会说,尽管生态位构建“可以影响甚至导致自然选择的演化过程”;但“生态位构建自身并非演化过程,最多就是一种变化的环境”。这种观点有所欠缺:更一般地将生态位构建与环境变化区分开的是,生态位构建以有序、有针对性与持续的方式改变了自然选择。生态位构建不仅仅会改变自然选择,而且还会通过对选择的方向和强度施加统计偏差来指导自适应演化。生物对适应性景观进行设计。例如,对美国东北部15个森林中原生植物群落上生物入侵进行的分析显示出惊人的一致模式。尽管蚯蚓种类和丰度、本地植物组成与特性、以及地理区域存在差异,但在所有地点都检测到了与蚯蚓相关的相同反应。
如Lewontin所建议的那样,相比于将演化视为适应性景观,将其视为生物走过蹦床的过程可能会更好。但是,已有示例显示,随着生物在演化过程中的变迁,它们不仅会抑制自身的适应能力,而且还可以改善环境,从而改善其适应能力。该过程强大到可以影响演化结果的确定。例如,蚯蚓并没有上升到典型的陆生动物生理的适应高峰。它们反而又进行了一次演化旅程,在陆地到达了一个难以置信的水生动物生理适应高峰。
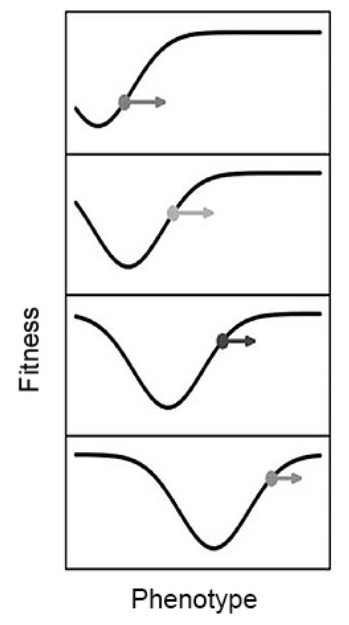
6. 相互作用的地形图
6. 相互作用的地形图
最近,Mark Tanaka、Peter Godfrey-Smith和Benjamin Kerr——一个理论生物学家与哲学家组合的团队——设计了一种新颖的数学框架[26],用于对自然选择和生态位构建进行建模。他们以两个关联的地形共同描述适应性演化,而不是仅用单一的适应性曲面:有机体通过在适应性(adaptive)景观上攀登而演化,同时环境的演化是通过在构建性(constructive)景观上攀登而发生的。两者之间的关系决定了特定生物的成功程度。就好像不追踪雌孔雀择偶偏好的演变,就无法充分了解雄孔雀尾巴这类第二性征的演化。因此,如果不考虑生态位的构建,就无法描述适应性随生物对环境特征的改变所产生的演变。Tanaka和同事使用他们的生态位构建演化框架来分析三种微生物演化案例的动力学,其中微生物参与了不同形式的生态位构建。
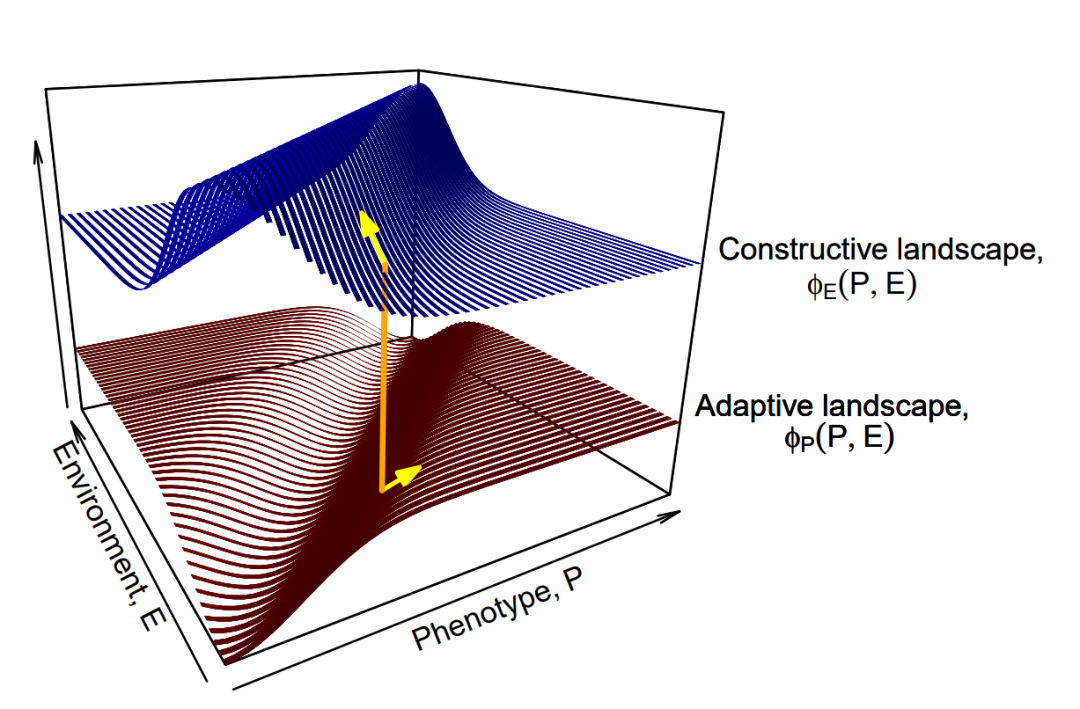
所谓的费舍尔基本定理(Fisher’s fundamental theorem)描述了自然选择推动种群向改变自身来适应地形的方式:通过自然选择演化的种群适应性的增长率等于适应性的遗传方差(即导致生存及繁衍能力产生变化的遗传变异量),从而导致生存和繁殖差异。统计学家兼遗传学家罗纳德·费舍尔(Ronald Fisher)认为,由于种群适应性无法无限增长,因此必须考虑环境的恶化来平衡这一过程。Tanaka和他的同事们构建的地形耦合模型从数学上表明,在生态位构建中,这种适应性的守恒不一定存在。生物可以通过改变其环境的方式来提高其绝对适应性(如在山峰筑丘),同时也可以破坏其环境并降低其适应性(如挖坑)。
该模型和解释了蚯蚓与其他重塑环境的物种(包括人类)的演化有关。欧洲蚯蚓在北美森林中取得的巨大成功,很可能不是自然选择使其适应森林环境的表现,而是它们改变森林生态以使森林更适应自身能力的结果。对于人类而言,这一点也适用。驯化植物、耕种土壤、采用新的灌溉方法和施用肥料——它们都增加了土地供养人类的能力。通过反复改善环境,我们已经提高了我们的绝对适应能力,这就好似我们在适应性景观上建造了越来越高的塔,然后爬上塔来到达地形的巅峰。
7. 性状演化:自然选择+生态遗产
7. 性状演化:自然选择+生态遗产
演化生态学家Tobias Uller 与 Heikki Helanterä设计了一种方法,将生态位构建结合到另一个普遍的演化模型中,即以群体遗传学家乔治·普莱斯(George Price)命名的普莱斯方程[27]。该模型建立在两个核心的演化过程之上:对性状的自然选择随时间的演化;以及性状在几代之间有偏好性的传递(或继承)。这些过程中的每一步都可以进行统计定义和测量,从而提供一种计算性状随时间变化(即演化)以及选择或传递贡献量的方法。自然选择项由性状的可遗传性(遗传差异导致性状差异的程度)与选择差异(衡量性状对适应性的影响程度)的乘积给出。例如,如果大型狗相比小型狗拥有更多的幼狗,但是耳朵直立的狗与垂耳的狗后代数量相似,那么体型大小会具有较大的选择差异,而耳朵形状的选择差异将会很小。感兴趣的性状可以是任何事物,包括对环境的适应程度。由于遗传力与遗传变异的数量有关,因此当关注的性状是适应性时,普莱斯方程中的自然选择项广义上与费舍尔基本定理等价。
相比之下,性状传递项代表的是自然选择之外的其他因素,这些因素会影响性状随时间的变化,包括其对遗传何种性状的影响。有趣的是,Price将其称为“环境变化项”,这似乎特别适合于此类场景。蚯蚓并不是随意地改变土壤——有时会给后代留下一个更好的世界,有时会变得更糟,它们也没有持续分解土壤。相反,蚯蚓一代又一代地提高了土壤的质量,通过生态位的构建提高了自身的适应能力,并为其后代留下了有偏好性的生态遗产。正如费舍尔(Fisher)认为环境应该恶化的假设一样,历史上,当关注的性状为适应性时,研究人员倾向于认为普莱斯方程中传递偏差项为负值。这是因为突变通常对生物不利,也因为环境影响被(错误地)认为不会被遗传。结果,与传递项有关的过程在演化中所起的作用相当程度上已被忽略。当有机体为其后代构建或选择更好的环境时,即使没有自然选择,它们也可能引起适应性演化[28]。
Uller和Helanterä使用普莱斯方程指出了生态位构建可以影响演化的其他方式[29]。在演化分析中,可遗传性(由于遗传差异而导致的性状差异的多少)通常是通过统计(例如,父辈与子辈间的回归)来估计的,该统计数据比较了父母和后代的性状相似性(例如,大狗的幼犬是否更大?)。父辈将改造后的环境传递给后代,使得可遗传性的估计值偏大。
对该可遗传性估值偏大的现象,蚯蚓又一次提供了有用的例证——这次是是粘液——因为它们分泌粘液以保护其皮肤外层,并且粘液的分泌对土壤特性十分敏感[30]。在蚯蚓处理程度不同的土壤条件下进行测量,蚯蚓父辈与子辈之间的粘液水平之间存在很强的相关性。但是,这并不是因为蚯蚓会将“多余的粘液”基因传给婴儿。亲子后代的相似性和高遗传力的出现主要是由于生态学而不是遗传遗传:后代“继承”了其父母的土壤(物质环境与微生物组),因此产生了类似数量的粘液。如果研究人员调查粘液水平如何对入侵的蚯蚓进行自然选择,那么他们很可能会检测到性状发生强烈变化,这意味着选择低量的粘液分泌。但实际上,这是生态位构建的生态效应,因为蚯蚓亲本已经为后代改善了土壤环境。目前尚无对自然选择的遗传反应,但性状发生了系统性和方向性变化,表明性状与环境适应性密切相关。
8. 适应性跷跷板
8. 适应性跷跷板
生态位构建对演化的另一种影响方式是改变性状影响适应性的方式(即改变选择差异,这是普莱斯方程中自然选择项的一部分)。例如,对于蚯蚓遇到贫瘠土壤及食物缺乏倾向于迁徙的性状[31],实验研究表明,可以通过改善栖息地质量来减少自身(和其他蚯蚓物种)迁徙的需求[32]。它们没有迁移到更富裕的环境中——而是灌溉、施肥和重新种植土地,将其转变为优质土壤。
现在考虑在入侵的蚯蚓种群中,存在“快速”和“慢速”两种扩散者。拥有“快速扩散”基因的蚯蚓可能会定居在土壤中,但由于原始土壤的质量较差,因此最初适应性较低。但随着它们努力改善环境,更多的无脊椎物种以及有益的侵入性灌木和动物(对它们有益)被吸引,因此它们的适应性进一步提高。最终,蚯蚓种群变得密集,以至于其适应性因竞争和资源过度开发而下降,从而引发了新一轮的传播浪潮。相比之下,拥有“慢速扩散”基因的蚯蚓最初比定居者具有更高的相对适应性。但是,随着被入侵森林土壤质量的改善,慢速扩散的蚯蚓相对适应性会下降,直到它们过度分散为止。在生态位构建和竞争的推动下,快速和缓慢的扩散者陷入了周期性的战斗。回到适应性景观的比喻,这里的地形就像一个跷跷板:慢速扩散受到青睐,但随着快速扩散者改善土壤,跷跷板向另一端倾斜,而快速扩散则成为适应性时的最高点,直到跷跷板再次翻转。
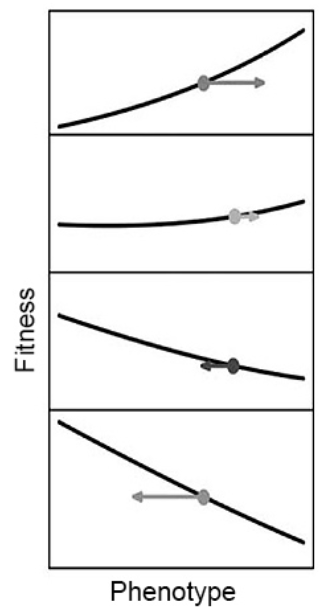
所有这些形式化的数学分析都有助于加深对生物改变环境后果的理解。为了理解适应性演化,研究人员不仅需要了解生态位构建如何通过自然选择而演化,还需要了解生态位构建如何改变自然选择的环境来源。我们需要扩展适应性景观的比喻,以刻画由生物所引导的环境变化。不断演化的种群不像僵尸登山者那样盲目地攀登适应性高峰,而是更像勤劳的地形设计师。它们配备了挖掘和建造设备,可以根据自己的需求对地形进行重塑。在人类生态位构建和生态遗产正在破坏地球的生态并推动人口爆炸增长的现状下,了解生物体如何通过重构生态而提高适应性变得更加紧迫。
原文链接:
https://aeon.co/essays/organisms-are-not-passive-recipients-of-evolutionary-forces
参考资料
(参考文献可上下滑动查看)
复杂科学最新论文
集智斑图顶刊论文速递栏目上线以来,持续收录来自Nature、Science等顶刊的最新论文,追踪复杂系统、网络科学、计算社会科学等领域的前沿进展。现在正式推出订阅功能,每周通过微信服务号「集智斑图」推送论文信息。扫描下方二维码即可一键订阅:

推荐阅读
点击“阅读原文”,追踪复杂科学顶刊论文
微信扫一扫,分享到朋友圈











