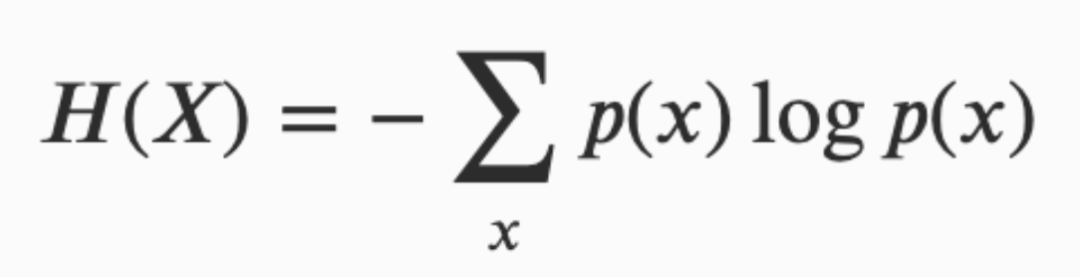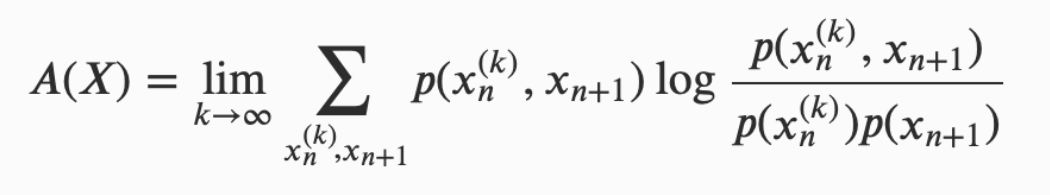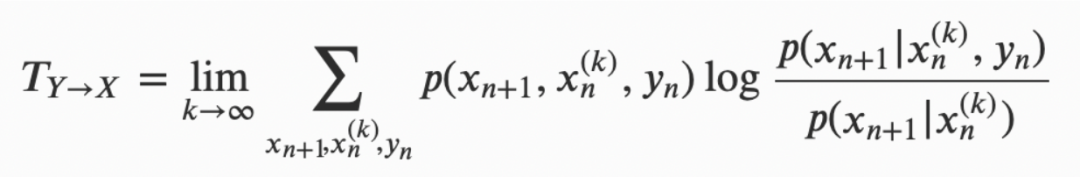生命是什么?这一追问至今仍在进行中,目前的答案以定性居多。而随着在自然界发现越来越多新型生命形态,以及新技术所创造的许多类生命系统,生命系统与非生命系统之间的明确边界正在不断受到质疑。近日发表在Theory in Biosciences杂志的一项研究,用信息论分析生命系统与非生命系统在信息架构上的区别,并讨论量化构建“生命光谱”的可能性。论文通讯作者Sara I. Walker任职于亚利桑那州立大学和圣塔菲研究所,从事复杂系统和生命起源、天体生物学研究。
Hyunju Kim, Gabriele Valentini, Jake Hanson & Sara Imari Walker | 作者
十三维 | 译者
集群行为(Collective behavior)被广泛视为生命和智能系统的标志属性。但在许多已知的无生命纯物理系统中,也有表现出集群行为的例子。这导致人们倾向于接受使用简单物理模型来解释生命的集群行为。为理解发生在生命实例中的真实情况,重要的是确定生命和非生命系统的集群行为之间,是否存在根本性的差异,以及从简单的物理实例建立对生命现象解释时直觉的局限。在这里,我们提出了一个框架,将生命和非生命集群作为基于各自信息架构的连续体进行比较:即信息究竟是如何跨越不同自由度进行存储和处理的?我们回顾了从信息论角度描述集群现象的各种例子,并对未来基于信息架构量化生命集群行为的方向提出了建议。
被我们冠称为”生命”的这一大类现象,迄今仍无法进行具体的科学定性。虽然此前已经提出了许多关于生命的定义(Clel& & Chyba 2002; Tirard et al. 2010; Benner 2010; Walker & Davies 2013),但几乎都是描述性而非定量的。人工生命和系统化学方法正在推动我们越来越接近在实验室中实现”类生命”(life-like)系统。基于已经发现的、几十颗围绕在其它恒星运转的宜居星球,人类在地外世界寻找生命的努力也在加快步伐(Kopparapu et al. 2014; Dressing & Charbonneau 2015; Anglada-Escudé et al. 2012, 2016; Gillon et al. 2017),一些新近投资的航天项目,例如龙飞船(Dragonfly),甚至即将访问土卫六去搜寻地外生命的痕迹。
在没有量化指标确定我们发现的东西是否真是”生命”的情况下,我们寻找生命新样例的努力很大程度上是盲目的。因此制定严格、量化的方法来描述生命是什么,变得越来越迫切。以往大多数努力都采用现象学方法来定义生命最重要的属性,例如将复制、新陈代谢活动、进化能力、细胞边界等作为关键属性(Clel& & Chyba 2002)。鉴于寻找生命最终还是需要客观标准来对存在证据进行评估,因此有必要采用新的、定量的方法来理解和测量一个系统是否是“活的”,以及若是活的则哪些因素是必需的。这项任务的根本挑战在于,我们能用什么来度量?
在众多可以为定量方法提供基础的属性中,生命系统存储和处理信息的能力有潜力成为最基本和最独特的属性(Szathmáry 1989; Küppers 1990; Yockey 2005; Davies & Walker 2016)。生命通过操控信息来重塑物质结构并使其发挥功能的能力,与其它任何种类物理系统中看到的情况都非常不同。虽然这种观点确实在各种流派中越来越受到青睐,但它仍然存在很多问题有待说明:我们可以给生命系统赋予什么可度量的指标?它会是信息论的吗?会基于信息流下的网络结构吗?抑或只是一种全新的形式主义?
为解决这些问题,重要的是现在就开始用我们目前手头的工具来重构“生命是什么”的问题,即使随着理论和实验的迭代进展,这些工具最终可能会被更细致的量化数学理论所取代。发展任何理论都应该有可验证的结果,相关可观测物要有明确的定义。对比下其中最有希望的数学工具,就是信息论和网络科学的工具,以及最近对信息理论的扩展。这些工具能允许评估因果交互作用;此外迄今为止,还没有其它任何方法有机会完成量化一般生命过程的使命。
任何科学方法若想在诸如量化生命及其特性这样困难的领域取得进展,必将遭致严重的批评和挑战。例如,人们可能反对专注于生命信息的一面,认为信息论方法不仅适用细胞内发生的过程,同时也能用于描述社会和城市过程,范围过于宽泛将导致定义不清。此外,从信息方法中得出的结论也受测量样本数的限制,并且并没有一个有效方案来解决这个问题。
但我们认为,这两方面都是优点而非缺点。没有理由认为”生命”只是一种在天体生物学假设下、仅发生在化学中的现象。尤其是在人工生命或复杂系统领域中,前面假设并不重要。我们反而认为化学仅仅是相关的、只是生命涌现的物理学尺度,而生命本身是一个更广泛的现象,会在不同尺度上反复出现:从化学到细胞,再到社会——它会普遍地涉及信息(抽象属性)与物质间的交互作用。若确实如此的话,这就给我们提供了一个契机,使我们能在仅有一个生物圈的有限样本内,也能研究各种跨越不同尺度的生命实例(Walker 2017)。我们期望这种普遍性方法不仅能阐明地球上不同系统和尺度的生命结构,也能揭示其它生命系统的可能性——即有哪些别的物理系统也支持与已知生命实例具有相同的信息结构。
在本文中,我们提出了这样的观点:在有生命系统和无生命的系统之间可能没有明确的、非黑即白的区别,二者之间不存在鸿沟和界限。执着于生命/非生命的二元对立一直在阻碍着研究进步,相反,顺着如图1那样可能存在状态渐变的”生命光谱”,可以看到,即使复杂程度相似,有些系统也比其他系统更有生命力(alive)。而要衡量“生命力”(aliveness),信息就是一种很好的度量,毕竟它与尺度无关,且集群系统向更有生命力系统的过渡也可能需要信息。对此测量可以集中于给定物理系统中可能发生的转变:更有“生命力”的系统允许更多可能的转变,并经常促成更多不可能的转变。一个例子是,利用农业信息的农耕活动如何改造了我们人类的文明,导致产生了改造物质的能力,而这些能力在这一革新之前是不存在的;或者人文学科对科学的发明如何导致人类物理上实现从地球发射到太空的能力,显然这在万有引力定律的知识之前也是不可能的(Walker 2016)。在这方面,生命可以说是在探索足以导致事物发生的相邻可能性(adjacent possible,Kauffman 2019)。其中转变可能性的数量与生命物质的信息结构有着深刻的联系,因为只有当生命系统获得了物理世界的信息,它们才可能控制与这些信息相关的过程:更多的信息意味着对更多自由度的控制。
图1:生命光谱中的信息。该图阐明了一个如何将生命系统视为同一物理现象不同层级例子的概念框架,关键在于信息结构的区别。以这样的尺度上对生命系统进行排序是否可能;以及若可能,哪种度量更能刻画这个尺度的特点,目前仍是一个悬而未决的问题。图中复杂系统分别代表:一组非生物的化学化合物、细胞内的生化反应、由多达5万个细胞组成的团藻球形种群、蚂蚁群体的集群行为,以及嵌入城市的社会结构。这当然并不全面,但说明了代表不同”生命力”程度的系统在不同尺度缩放下可能的样子
生命可能存在一个光谱的想法是信息方法的自然结果:如果信息蕴含的某种概念确实是生命物质最基本属性的基础,那么生命就应该能用其信息结构和因果结构来量化(Walker & Davies,2013)。然而,作为围绕生命属性的信息和因果结构,一旦延伸到纯物理系统,就开启了统一描述生命和非生命系统并量化它们相似和差异的可能性。当我们把目光投向行恒星和星系等大质量天体来研究引力物理学时,就像承认引力是宇宙的一个普遍属性一样,我们应该认识到信息也是,而生命过程就是研究信息物理学最典型的例子。
可见重要的是,任何量化生命的尝试都必须能处理非生命模仿生命系统行为的情况。因此在这篇文章中我们关注集群行为——它被广泛认为是生命的标志属性,同时又能在许多非生命系统中观察到。事实上简单的物理模型经常被用来模拟生命的集群行为,这就引出了它们是否存在根本差异的问题。我们认为,通过研究集群中的信息流,将获得信息理论是否能够区分生命和非生命系统的深刻见解:假设两个系统,通过不同机制表现出相同的聚集行为(aggregate behavior),一个是活的而另一个不是,信息论就能发现这些差异。信息论关注的是捕捉空间和时间相关的结构,因此可以使用信息论度量来确定两个系统是否使用了相同的规则集合。我们的假设是,生命能通过各种相关操纵(例如,对自身状态的干预)主动控制自身的状态,而非生命则不能。因此信息理论将成为辨别非生命和生命集群相关性结构差异的有用工具。就此而言,生命过程的光谱可能由控制(因果)和信息结构的组合决定,组合沿着从生命力较少到生命力较多的频谱对各种实体进行量化。虽然”生命光谱”的精确度量和尺度仍有待确定,但我们将在本文概述一些可能进一步有用的关键思想。
信息是一个抽象的概念。虽然由克劳德·香农(Claude Shannon)开创的信息数学理论将某些方面形式化了,但它并非对“信息是什么”的完整描述。尤其,我们并没有一个具体的框架来理解对信息之于物理究竟是什么。信息可以在不同的物理介质之间复制,这意味着它并不是严格意义上只与物理质料相联系的物质属性。例如,信息可以从作者的头脑中复制到这一页文字上(通过写在电脑),或者复制到印刷的纸张上,并最终按照预期功能复制到读者的大脑中(例如这里你阅读到的)。这些事件在空间和时间上是分离的,并且发生在完全不同的物理介质上。就像许多信息传输的例子一样,这表明信息不可能仅仅是大脑的化学湿件、或计算机处理芯片上的一种属性。正是在这个意义上,信息才是“抽象的”(同样人们也可以认为“能量”是抽象的,因为它能在不同的物理系统之间流动,并以不同的方式储存或使用——如化学能、机械能等。但相比之下,我们确实对什么是物理上的能量有一个非常清晰的认识)。
然而,信息也必然是物理的。为了使信息存在,它必须在物理自由度中实例化(L&auer et al. 1991),因此信息的动力学取决于物理自由度下动力学。在生物学中,信息的作用更为突出,它似乎具有“自己的生命”(Davies&Walker 2016),对生物过程的解释暗示“信息”本身就具有因果效应(Davies 2011)。生命拥有能够存储和处理信息的惊人能力,不仅对于理解生命本身的起源(Yockey 2005;Walker&Davies 2013),对于理解生命系统如何在我们观察到的时空尺度中组织起来,也是一个具有核心意义的课题。信息传递和信息处理通常被认为是集群行为背后的驱动力,例如蚂蚁和蜜蜂的寻巢行为(house-hunting)、椋鸟群和鱼群的奇妙运动、人类群体中的车道形成等等,直到细胞群的水平(Franks et al. 2002;Couzin,2009;Deisboeck&Couzin,2009;Moussaid et al. 2009;Couzin et al. 2011)。然而,虽然已被大量实验和分析研究过,以上这些集群的信息机制却往往只以生物体的具体物理量为特征、仅以非形式化方式就被定义了。
为超越单纯的描述性分析,或具体的案例研究,有必要引入形式化信息内容和信息流的方法。物质属性已经被量化了几个世纪,信息则相对是更新的进展,它始于香农的开创性工作。在信息论中最基本的量称为熵,H(x) 被定义为(Shannon 1948):
其中 p(x) 是随机变量 X 处于状态 x 的概率。当 p(x) 是所有可能状态的均匀分布时,H(X) 是最大化的,它通常称为香农熵。香农熵可以被描述为你在得知一个事件结果时可能体验到的惊讶程度:事件越不可能发生,你在得知事件时就感觉越惊讶。从这个角度来看,信息的概念与减少不确定性密切相关,因为一个事件的不确定性越小,你对事件结果的惊讶程度就越小,它所包含的信息量也就越少。香农熵构成了信息论的基础,其它大多数信息概念和度量都是基于香农所发展的一般信息概念的基础上衍生出来的。然而仅仅用熵并不能完全体现”信息”的概念,因为信息还需要发送者和接收者(Adami 2016)。在这方面,信息被定义为给定其它过程下某个随机过程不确定性的降低,并由所考虑过程间共享的熵的大小来表示(Cover & Thomas 2005),这被量化为互信息(mutual information):
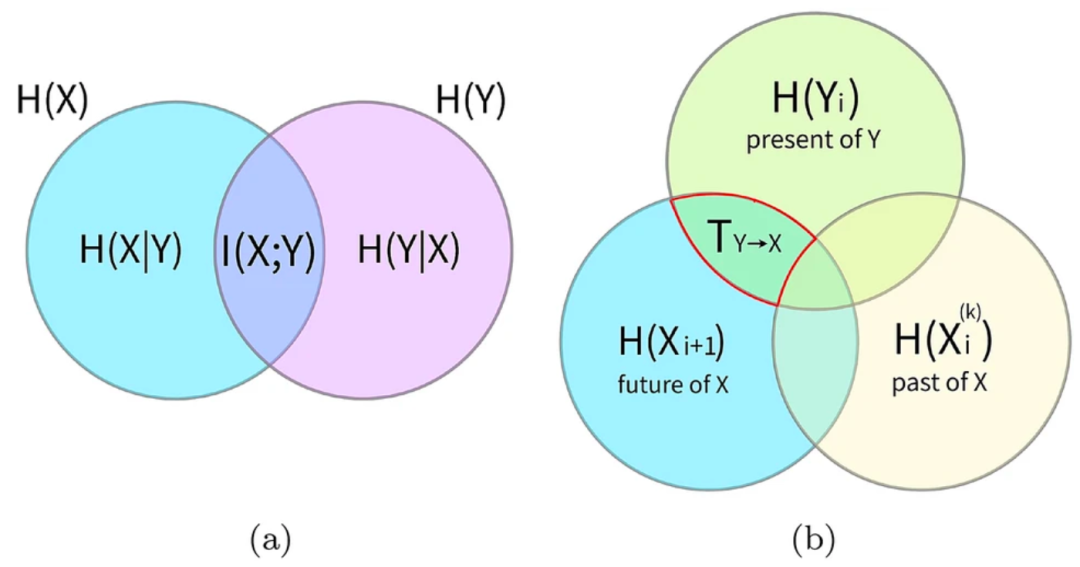
互信息可以很方便用维恩图来可视化,即每个过程的熵相互重叠的区域(图2a)。当考虑某个过程的熵,去掉与其它所有过程的互信息,我们就得到了条件熵:即了解其它所有过程知识时,该过程还剩下的不确定性。因此,互信息度量的是我们从所有其它过程知识中获得有关某个过程的信息。
互信息也被用来量化存储在一个系统和/与它各个组成部分中的信息量。最常用的测量方法是超额熵(excess entropy)和主动信息(active information)。超额熵(Crutchfield & Feldman 2003)被定义为一个过程过去和未来之间的互信息,衡量某个过程可以通过观察其过去的行为来解释的未来的不确定性;主动信息 A(X) 也提供了类似的度量,但它不考虑整个过程的未来,而只关注和预测过程中即将抵达的下一个状态(Lizier et al. 2012a):
(其中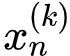 代表 X 中从时间点 n 开始过去 k 个状态:{x𝑛-𝑘+1, …, x𝑛-1, x𝑛}。)
代表 X 中从时间点 n 开始过去 k 个状态:{x𝑛-𝑘+1, …, x𝑛-1, x𝑛}。)
虽然互信息允许定义和量化信息,但它只是一个对称的量,不能捕捉到随机过程之间的方向性关系(如信息传递)。为克服这一信息量化处理的局限性,Schreiber(2000)引入了传递熵(transfer entropy)的概念,它是给定某个随机过程过去(如历史信息)和一个或多个其它过程当前知识了解下,对该过程未来状态的不确定性减少的一种度量(Schreiber 2000; Kaiser & Schreiber 2002):
其维恩图表示如图2。传递熵利用时间方向性来克服互信息对称性的限制,它是对两个或多个过程之间的定向传递的预测信息的度量(Lizier&&Prokopenko 2010),因此也不一定就意味着因果作用(James et al. 2016);但它的逐点变量(point-wise,或局部变量)可以为集群行为的时空动力学提供有用的洞见(Lizier et al. 2008b)。
有关信息在生物过程中的作用,以及生物学是否需要一种不同类型信息的形式化(在现代信息论中通常被称为功能信息或语义信息),一直存在着长期争论 [例如, Yockey (2005) & Godfrey-Smith & Sterelny(2007)]。我们认为,目前阐述的信息理论,特别是考虑到分布相关性(Schreiber 2000)和因果信息(Hoel et al. 2013; Ay & Polani 2008)测量方面的最新研究,足以在信息如何构成生命的问题上取得进展。
具体而言,我们认为生命系统最与众不同之处是它们的因果结构和信息架构(Walker 2017)。现代信息测量方法,如传递熵(Schreiber 2000)、因果信息流(Ay&Polani 2008)、有效信息(Hoel et al. 2013)、因果特异性(Griffiths et al. 2015)、整合信息(Oizumi et al. 2014)和整合时空模式(Polani et al. 2016)等等,所有这些都有希望从信息论角度对生物过程提供多样而丰富的见解,使我们有能力了解如何在各种时空尺度上描述生命的信息架构和因果结构。
例如,速度和方向信息通常是在鱼群(H&egard et al. 2012)和椋鸟群(Cavagna et al. 2010)协调运动的背景下研究的物理量,此类信息在群体中的传播已经用统计物理工具进行了建模(Bialek et al. 2012),各种行为机制 [如群感响应(quorum response)——许多动物采用的一种共识机制(Pratt et al. 2002;Wardal et al. 2008;Sumpter&Pratt,2009)] ——也进行了类似的研究。这些机制允许集体中的个体建立反馈回路,以放大或抑制信息的传递(Couzin 2009)。尽管研究这些群体信息传递、存储和处理的形式化方法还不太成熟(Dall et al. 2005),但对这些现象的大量关注为形式化应用方法和发展新理论提供了有力依据。
在下面内容中,我们并不认为存在一种量化生物集群行为”正确的信息度量”立场,而是主张在适当的控制下,通过在不同的生物数据集上应用多样化的测量方法,解决生物系统如何在空间和时间上操纵因果相关结构以执行功能。我们会通过解析生物系统区别于非生物系统的信息架构(Walker et al. 2016),并希望利用这些见解构建解释“生命是什么”的新理论。
形式上就我们旨在本节中提出的意义而言,集群行为的信息结构(以生命作为旨趣实例)是相对未被探索的开放的研究领域。尽管如此,一些研究已经概述了它的一些显著特征。例如,Danchin 等(2004)将个体可获得的信息分成个体信息(personal information)和社会信息(social information),前者通过与环境的直接互动获得的,后者则受益于其他同伴的行为获得。进一步又可以将社会信息分为通过信号传递(signaling)和间接观察(indirect observation)获得的社会线索或公共信息。信号传递是有意传递的信息,而社会线索(如觅食者的位置揭示了食物来源)和公共信息(如觅食者的表现提供了食物来源质量的信息)则缺乏此类特征。在集群行为背景下,Moussaid等(2009)强调了直接信息传递和间接信息传递的区别。按照 Danchin等(2004)的说法,直接信息传递对应明确的信号传递以及对社会线索和公共信息的间接观察,间接信息传递则是个人信息的一个子集(Danchin et al. 2004),它只包括个体之间通过环境中介传递的信息。间接信息传递通常被称为共识自主性(stigmergy),它包含了一种沟通形式,在这种形式中环境的功能是一块共享的黑板,个人可以修改黑板来书写信息,也可以感知阅读信息(Grassé 1959)。间接信息传递最突出的例子无疑是某些种类蚂蚁的信息素铺设和跟随行为(Goss et al. 1989;Hölldobler & Wilson 1990)。
然而上述定义,主要关注的是个体获取与可能传递信息的不同方式。一个量化集群行为信息结构的形式框架,还应该考虑信息加工的其它基本方面。事实上,一个集群系统的内部计算不仅是组成部分之间信息传递的结果,还可能要求它们具有存储和转换信息的能力(Crutchfield 1994;Feldman et al. 2008)。这其中一些特征可能存在争议,例如目前还不清楚,在什么意义上,集体趋化作用(collective chemotaxis)会要求个体存储它们位置以外的信息,或可能只是要求它们做出反应——无论这是否是由进化编程存储的一组反应之一。
因此,集群行为信息架构的正式框架可以包含所有这些元素,重点在它们间的相互作用及随时间的演变。在这个方向上的第一步是接受一种独立于应用领域的通用语言,使我们有可能对信息处理的所有方面进行正式建模。当然这种共同语言要由信息论提供。
信息论度量方法引入后不久就首先被用于研究集群行为:推断蜜蜂的摇摆舞(Haldane & Spurway 1954)和蚂蚁信息素踪迹(Wilson,1962)所编码食物来源的方向信息大小。但应用信息论对集群现象进行系统研究却相当零散(Dall et al. 2005) 。例如,统计物理学将互信息用于研究二维伊辛模型(Ising model)中的相变(Matsuda et al. 1996;Gu et al. 2007),由维切克模型 [1](Vicsek model)驱动的自推进粒子蜂群模拟(Wicks al.,2007),以及调节模型(regulatory models)中的随机布尔网络(Ribeiro et al. 2008)。在所有这些情况下,互信息在无序或接近相变阶段时均达到峰值。互信息和块熵(block-entropy)——一种测量有限连续事件不确定性熵的变体(Shannon 1948),已被用于研究蚂蚁信息素铺设模拟中的群体决策(Klyubin et al. 2004),此项研究表明有限的噪声有利于信息传递(Meyer 2017)。最近,Gelblum 等人(2015)的研究则显示,附着在集体运输上的蚂蚁会向系统输入信息,但这种信息只在短时间内有效。以上这些测量方法不仅被用作分析工具,还被应用于人工进化控制器,设计模块化的集群运动行为(Prokopenko et al. 2006)和多机器人系统(Sperati al et al. 2008)等等。
信息论涉及大量不同科技领域信息的量化、存储和传递(Cover & Thomas 2005)。例如在生物学的背景下,信息论被广泛用于研究神经系统的功能并确定相关脑区结构(Honey et al. 2007;Vakorin et al. 2009;Nigam et al. 2016;Ito et al. 2011;Lizier et al. 2011;Vicente et al. 2011)。主动信息(见前面公式)被用于研究元胞自动机的信息存储(Lizier et al. 2012b)、神经信息处理(Wibral et al. 2014)以及蜂群动力学(Wang et al. 2012)等,而超额熵则主要应用于复杂物理现象的研究(Crutchfield&Feldman 2003)。
虽然其它信息流和因果信息度量方法的应用将为我们讨论的问题提供更多见解,但这里我们主要会关注传递熵的信息处理的方法。并在后面为所选的传递熵提供了一个说明性的示例,驱动来自大量不同领域传递熵的应用,如神经科学和金融(Bossomaier et al. 2016),包括互信息及传递熵都已被用于研究二维伊辛模型不同阶段的信息流(Barnett et al. 2013)。Lizier 等(2008b)用它来研究元胞自动机,并表明质点(particles,定义为滑翔机(gliders)和域壁(domain walls))是这类模型中传递信息的主要手段。
Wang 等(2012)研究了人工粒子群体运动模拟中的信息级联,表明它们以波浪的形式在群中荡漾。在模拟中使用传递熵框架研究的其它集群行为还包括酵母细胞周期的动态调节网络(Kim et al. 2015;Walker et al. 2016)(见下文讨论)和多代理系统中的共识实现(Valentini et al 2018)。此外在动物集群行为研究中也得到了一些应用。例如,研究成对和小群的斑马鱼的信息传递,为识别领导关系提供了有用的工具(Butail et al. 2014,2016;Mwaffo et al. 2017),以及鱼类U型转弯过程中的信息和错误信息交互作用(Crosato et al. 2018);还有研究蝙蝠(Orange & Abaid 2015)或蚂蚁和白蚁的引导者-追随者关系(Valentini et al. 2020),粘液霉中的信息传递(Ray et al. 2019),以及成对鱼群之间的捕食者-猎物关系(Hu et al. 2015)。
既然不同度量指标如此宽泛且还都具有广泛适用性,对在不同系统中应用信息理论见解的最佳方法、甚至对这种方法是否可能,目前都尚无明确的共识,每种度量都提供了一种动力系统因果与相关结构在较低维度上的投影,因此并不能捕捉系统全貌。而将不同度量方法结合在一起,可以提供对生命和非生命系统结构更深的洞察,进而可以利用这些度量方法采取下一步行动,再发展出新的理论和度量方法,以了解生命究竟是什么。
集群系统经常面临需要做出集体层面共同协商的决定。这种决定是由大量的代理体作出的,往往遵循简单的互动机制:收集、传递和处理集体决定所需要的信息。群体决策过程中的信息传递最重要:没有信息的交流,集体之间就无法达成共识。尽管我们目前对群体决策机制有很好地理解,但个体规则在群体决策信息传递中的贡献却还鲜为人知。
为了解决这个问题,更好理解不同决策机制的信息状况,我们研究了源自同一决策问题(Valentini et et al. 2018)信息传递应用情况下的不同决策策略:从两个选项中价值较高的一个达成共识(Valentini et al. 2017)。我们考虑了两种决策机制,多数决规则(majority-rule)和选举模型(voter model),均通过集体中100个代理人产生意见。在我们的模型中(Valentini et al. 2016),代理人总是偏好两个选择中的一个。设其中地点 A 的质量为 1,地点 B ∈ {0.5,0.9}。代理人会在两个阶段交替行动:在探索期前往与当前意见相符的环境区域,对该选项质量进行采样;另外是根据估测的传播质量,在当地向共同地区其他代理人广播其意见的传播期。在传播期和探索期之间,代理人应用决策机制,并可能改变意见。当使用多数决规则时,代理人将把意见切换到大多数邻居所喜欢的意见(Valentini et al. 2015)。当使用选举模型时,代理人会简单地随机选择邻居的意见(Valentini et al. 2014)。在这两种情况下,我们都考虑了邻域构成,它会根据代理人的运动动态变化,但具有固定的基数(cardinality),通常一个代理会有5个邻居左右。
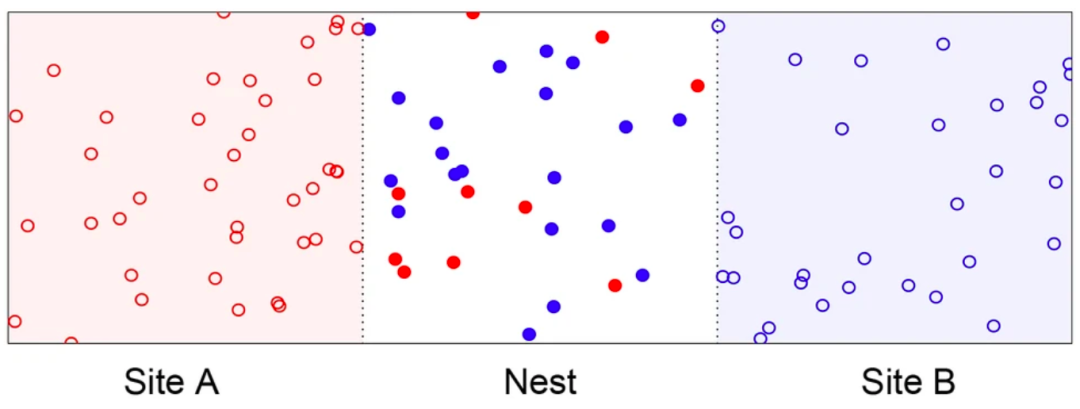
图3. 地点A(红色区域)、巢穴(白色区域)和地点B(蓝色区域)的模拟环境图示。实心圆代表处于传播状态的代理体,空圆圈代表处于探索状态的代理体。颜色代表选择的意见,红色表示选择 A,蓝色表示选择 B。图自 Valentini et al. 2018
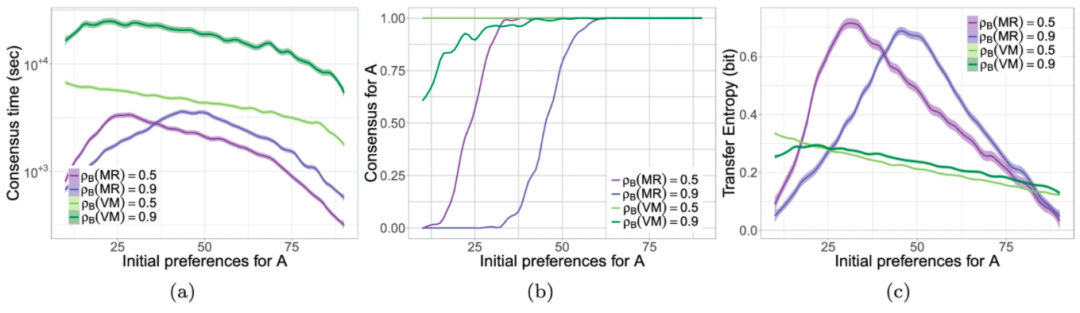
多数决规则和选举模型之间的机制差异导致了不同的表现:前者比后者更快(图4a),但准确度更低(图4b)(这一结果更深刻的讨论参见 Valentini et al. 2018)。
图4. a 共识时间(对数尺度),b 模拟收敛于地点A的共识比例,c 在N=100的群体中初始偏好地点A代理数(如 {10,11,…,90} )的传递熵。问题配置:多数决规则 𝜌𝐵(𝑀𝑅) ∈ {0.5,0.9},选举模型𝜌𝐵(𝑉𝑀) ∈ {0.5,0.9} 。数字报告了使用LOESS回归计算平滑条件均值和置信区间估计,数据跨度为0.1。图自 Valentini et al 2018
在此基础上,我们可以解释应用决策规则(即多数决规则和选举模型)的代理体从周围邻居那里接收的信息流。通过将传递熵应用于从空间多体模拟中收集的数据实现这点(见图3),我们发现,与速度和准确度一样,代理体之间传输的信息取决于决策机制。随着集体达成共识的决策时间增加,从邻居传递到应用决策规则的某个焦点代理体的信息量也会增加,并随着最终结果的不确定性宽松调整。这个例子突出了信息结构和局域规则之间的关系,即使在集群行为最终可能导致类似决策的情况下。它强调信息结构不仅是要做出决策的属性,同样是以各组成部分之间相关结构来说明微观动态和宏观尺度特征关系的一个属性。
为进一步说明这一点,可以使用我们中一些人的最近工作,将泰诺胸蚁(Temnothorax ants)纵队行进行为(tandem running behavior)的信息流结构与另外两种白蚁进行比较(Valentini et al. 2020),分别是精巢散白蚁(Reticulitermes speratus)和台湾乳白蚁(Coptotermes formosanus)。通过聚焦泰诺胸蚁的纵队行进行为,使用传递熵来研究引导者和跟随者蚂蚁之间的信息流。纵队行进允许对潜在地点知情的蚂蚁(引导者)引导不知情的蚂蚁(追随者)了解其获得的位置。纵队行进过程中,引导者会进行小段直线运动,跟随者则会进行更多种不同运动(如当记忆地标的时候)(Bowens et al. 2013),期间跟随者与引导者的身体接触会触发一系列重复动作。这种双向反馈(引导者引导跟随者跟随)被假设为实现了一种类似于人类教学的学习过程。与泰诺胸蚁不同,白蚁纵队行进的目的是确保新成对的雌雄白蚁夫妇在探索环境、寻找新巢址时不会失去对方。白蚁纵队行进的引导者并不知道潜在的巢址,因此并不试图将这些信息传递给跟随者。我们用传递熵分析了蚁群纵列对之间信息交流的幅度和方向,考虑了三种通信通道:运动、旋转和运动-旋转结合。根据相关信道的不同编码方案,对纵列对的空间连续轨迹进行了离散化处理:
我们发现,在胸蚁和白蚁的纵队行进中,引导者和追随者之间的信息交换存在以前未知的显著差异。在白蚁中,纵队行进的引导者操控着这对蚂蚁行为的所有方面(运动和旋转),而泰诺胸蚁却并非如此。与目前观点相反,我们的分析显示引导者和跟随者蚂蚁都扮演着不同的”领导”角色。从信息流的方向性看,虽然引导者指挥着蚁对的旋转模式,但决定蚁对运动模式的是跟随者。以这种方式,泰诺胸蚁的引导者-追随者组合通过不同的信道交换不同的信息,蚁对组合的交互作用为不同自由度的信息流编码了不同的方向。这是一个突出而明确的的例子,表明通过共享信息来进行群体决策是必要。因此对群体系统中信息流的严格量化有可能提供新的见解,揭示由于微观和宏观层面行为的微妙差异,新的计算类型(如决策)是如何从每个代理体的行动中产生的。
人们已经采用多种方法来刻画复杂系统,尤其是生物系统的因果结构特征。大多数人认为至少存在一些因果结构特性对生命过程的运行至关重要(Ellis et al. 2011; Griffiths et al. 2015; Auletta et al. 2008; Davies 2011; Roederer 2006)。为将生命系统的集群行为与非生命系统区分开,有必要了解信息架构与这些特征之间的关系。在这里我们提出,能够将生命系统的集群行为与其它复杂物理系统区分开来的,是信息架构和它物理实例的因果计算结构是否耦是合的。
在上一节,我们看到了一个通过改变局部交互规则来改变信息结构的例子。这里我们所说关注的因果结构,是指状态转换图的全局结构,它与系统的功能有关。在这方面,对相同的”行为”(状态转换图的因果结构)有多种可能的微观实现(Hanson & Walker 2019)。在本节中,我们将回顾以前有关复杂系统因果结构涌现特性的研究,如临界性、可控性和因果涌现,以展示它们与生物系统信息处理之间关系。我们的目标是具体讨论这些工作与因果结构在架构生物过程特有信息流中的作用,从而向量化生命的方向迈进。关于复杂系统和我们在这里涉及特定主题更广泛的讨论,请读者参考更普遍的研究(Munoz 2018; Liu & Barabási 2016; Oizumi et al. 2014)。
复杂系统往往表现出多种类型的集群行为,被称为相位(phases),这些彼此之间的行为与组成系统的单个实体行为往往截然不同,甚至大相径庭。当相变(phase transitions)发生时,即使系统一个微小的局部扰动,也可能导致全局状态从一个相变为完全不同的相(&erson 1972;Stanley 1971;Chaikin & Lubensky 2000)。大量文献都假设生命状态可能与某一类特定的相位相关,其含义是,生命状态的动态变化或非生命(随机)聚集态中生命的出现可以被描述为一种相变过程(&erson 1972;Hopfield 1994;Pollack & Clegg 2008)。
在这里,我们专注于文献中广泛讨论的两种不同动力系统系之间的相变:稳定和混沌,以及这两个区域之间平衡的临界系统。据研究从神经放电到细胞基因调控、动物运动的各种生物系统都是临界性的(Beggs 2007; Haldeman & Beggs 2005; Mora & Bialek 2011)。研究还表明,与临界点附近的系统相比,远离临界点的系统要么在有序阶段过于稳定而不能自适应,要么在混沌阶段过于不稳定而缺少鲁棒性。因此,生命系统的趋势是调整(例如通过进化选择)到临界点,这种临界动力机制被认为是内稳态和可进化性的驱动因素,以及生命过程的基本标志(Kauffman 1993)。
此外,复杂系统的计算能力,被定义为在执行某些任务时,内部逻辑结构输入和输出映射的数量和多样性,推测会在临界阶段最大化(Turing 1950; Crutchfield & Young 1988; Packard 1988)。这个猜想是作为与支持计算信息操作有关的问题而提出的,认为计算可能会在信息存储、传播和处理能力最大化的情况下(Kauffman 1993)更自然地在临界点(Langton 1990)附近发生。此外,Mitchell(2006)强调了通过网络系统中动态时间序列来理解信息动态的重要性。随后一些研究报告结果也表明,复杂网络中的信息处理在其临界阶段被最大化(Solé & Valverde 2001; Kinouchi & Copelli 2006; Ribeiro et al. 2008)。特别是 Lizier 等人(2008a)曾尝试改进以前这些计算特性的测量,对随机布尔网络(RBNs,Kauffman 1993)中的信息动态进行了彻底的定量研究。随机布尔网络被广泛接受为基因调控网络(GRNs)的模型,研究表明,量化存储和处理的主动信息和传递熵的平均值各自在临界点附近会达到最大值(Lizier et al. 2008a)。
从这里,我们可以看到一些典型物理模型局限性的暗示,例如平均场近似(mean-field approximations)和重整化群(renormalization group)方法,由于生物和非生物系统集群行为的差异,这些方法并不总是能转化为生物学所使用 [例如,向生命过渡过程的信息属性可能是不连续的,见 Walker & Davies (2013)]。也就是说,目前还不清楚平均场近似是否适用于包括生物学在内的一般情况,也不清楚通常应用于连续相变的重整化群是否适用于刻画生物相变的不连续性特征。解决方法之一是研究与生物功能相关临界系统的特征,以及这些特征如何表现出生物网络。例如,Daniels 等人(2018)最近证明了67个基因的调控网络是临界性的,而具有相似因果和信息架构的随机集合则不是临界性的,这显示了特定的因果和信息属性功能、与生物网络是如何通过其临界特性区分的。临界性甚至已被证明是随机基因网络模型中的一个可选择性质(Serra 2019)。
我们和其他小组最近的一些研究,强调了在与环境的因果互动和内部信息处理中适应性和鲁棒性之间的平衡(Walker et al. 2016),已知信息处理特性表现出从稳定阶段(以信息存储为主)到混沌阶段(以信息传递为主)之间临界转变距离的强烈依赖性。众所周知,临界系统具有生物系统的两个基本特征:对各种不同环境条件的适应性和鲁棒性。我们期望基因调控中的临界性普遍存在,这将为生物网络独特的信息处理和控制特性研究提供参考。
复杂系统研究的主旨之一是理解并理预测出系统涌现的全局动态,例如动态轨迹或最终状态(Liu & Barabási 2016)。这些代表系统集群行为的全局状态可以通过施加外部因果干预来改变。引导整个系统从初始状态到达期望的最终状态。这为许多领域应用都提供了强大的工具,包括从舆论动态相关的问题到流行病,从细胞的分化到社会性昆虫的巢穴选择问题等等。
最近的研究发展了复杂网络控制相关的数学基础,试图通过理解因果结构与各组成部分动态的更新规则来寻找严格的控制机制。Liu 等人(2011)发展了他们的前瞻性框架,将具有线性动态过程的有向复杂网络可控性问题简化为一个图论问题。利用他们的理论可以确定一组能够完全控制整个网络动态的驱动节点(driver nodes)。他们还发现,所确定的驱动节点往往是低度节点,并且总数主要由网络的度分布决定。
生物系统模型大多是非线性动力学系统,完全可控性既不可行也没有必要。相反,找到一种特定的控制机制,能够将一个给定的系统从特定的初始状态或轨迹引导到另一组期望状态或轨迹集合,是更为现实的,往往也足够的(Cornelius et al. 2013)。例如,控制基因调控网络的主要目的之一是将系统引导到特定的细胞状态,而非任意的表达水平。这种表型控制通常被定位在整个网络的一小部分中,以能驱动系统到期望吸引子的控制核来定义。不过,控制核的大小通常比一般随机网络还要更大(Liu et al. 2011; Kim et al. 2013; Gates & Rocha 2016; Choo et al. 2018)。这表明生物系统的可控性程度可能存在一定的范围,可以通过控制核的大小来量化。因此我们会将可控性程度视为生物系统信息架构的基本特征(Walker et al. 2016; Kim et al. 2015),以区别于所有非生物集群。
识别复杂系统中的因果关系是理解其涌现行为最基本的方法之一。然而这却是一项艰巨的任务:不仅因为复杂系统中的因果结构很复杂(坦率地讲它们确实很复杂),而且因为因果结构可以在多个时空尺度上进行分析,往往让我们搞不清楚哪种尺度是分析的首选(如果有的话)。长期以来,人们一直认为主因结构只能在最低(微观)尺度上定义,更高尺度上的因果结构是有用的,但只是对主因结构的粗略描述。即认为微观层面的因果关系确定了所有更高层次的因果结构,更高(宏观)层次的因果并没有实际上的因果贡献(Kim 1993,2000)。
这种观念下的还原论方法在科学各领域取得了广泛的成功,包括物理学和化学的重大发展。尽管如此,因果涌现的另一种可能——宏观尺度是因果结构的重要驱动,被一再提出来,因为它能对各种类型的生物集群行为提供更简单的解释:从表观遗传学到蚁群的群体决策,再到大脑中的心理状态等等。以往对这种因果涌现的研究大多局限于定性论证,而最近 Hoel 等人(2013)和 Hoel(2017)所做的定量研究表明,在一些系统中,因果结构可能无法完全被微观尺度结构捕获,相反宏观层面的因果结构会更有效、具有更丰富的信息。
如上所述,我们认为寻找一种复杂系统的控制机制,可以成为研究不同层次因果结构的有用工具,它能从局部(微观)层次因果深入洞悉驱动节点对全局(宏观)系统动态转向的影响。
如上节所讨论,在以往研究中已经暗示了生命系统信息架构与因果结构之间的关系。然而它们之间是否存在有意义的关系、若存在是否能够区分不同生物集群行为,这些问题却仍然是模糊、没有定论的。在这里我们提供了一个定量的例子,基于经验数据集建立了一个生物网络的动力模型,其规模较小故允许进行统计分析。由于系统生物学中高通量技术和数据驱动方法的出现,细胞生物通路的布尔网络模型是满足这一必要条件的最佳候选之一。
基于最近 Daniels 等人(2018)提供的迄今为止最全面的临界性调查,对从细胞集群数据库(Helikar et al. 2012)中获得的67个细胞调控布尔网络进行了研究,显示所有网络都接近临界状态。另一方面,Kim 等(2013)分析了各种生物分子的调控布尔网络,并确定了控制核——控制细胞网络达到预期状态所必需的最小节点集。研究表明,对于大多数网络而言,控制核只是网络组件的一小部分。因此,利用同类生物分子的调控布尔网络来研究生物系统、并将信息架构与其因果机制结合起来,是我们自然的选择。
在这里,我们以粟酒裂殖酵母(S.pombe,Fission Yeast)的细胞周期过程为案例,展示了如何在生物系统中研究信息架构与因果机制之间的关系(Walker al. 2016;Kim al. 2015),并作为生物中心功能体现的一个简单实例。
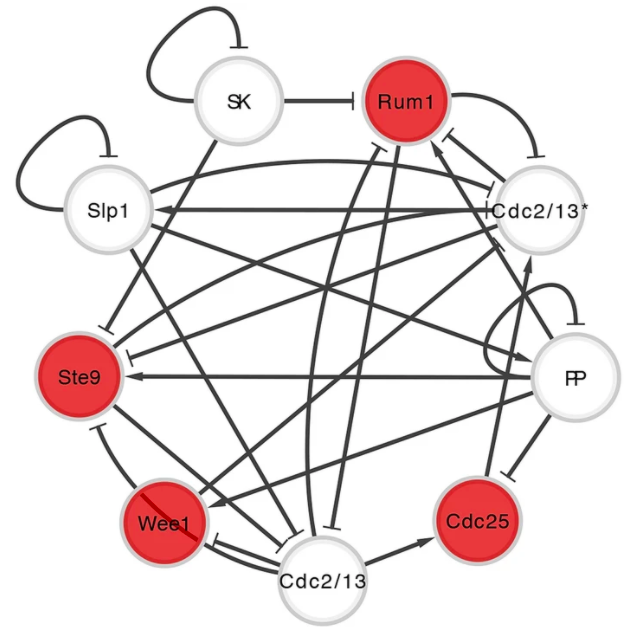
以布尔网络对裂殖酵母的细胞周期进行建模(图5),共有9个关键蛋白组成(Davidich & Bornholdt,2008),通过被称为 G1-S-G2-M 四个步骤的细胞功能进行控制:从细胞生长到DNA复制,再到细胞分裂成两个子细胞。在网络中,每个代表蛋白质的节点都有一个0或1的布尔值,分别表示其缺失或存在,每个节点状态的组合集被称为网络状态或网络配置。单个节点的状态,是与其它节点生化作用状态的函数,类型包括交互、抑制和激活。
将局部动力规则应用于每个节点,以模拟系统一个时间步长的演化。在对应细胞周期起点的特定网络配置上重复进行时间演化,从而以准确的顺序再现生物过程各个阶段序列。从512种可能的网络状态中的每一种开始生成网络状态空间,即所有可能时间演化的动力学轨迹,其景观就代表了裂变酵母细胞周期布尔网络的全局动态。网络状态空间包括13个吸引子、一个或一组多个网络状态,其中任意时间演化轨迹都会收敛。由于动力特性是确定性的,根据每个网络状态收敛到哪个吸引子,整个网络状态空间可以划分为13个不相交的子群。其中,约74%网络状态结束所在的主吸引子是唯一具有生物功能的吸引子(Davidich & Bornholdt 2008)。
Kim 等人(2013)最近关于控制这些生物调控布尔网络的时间演化轨迹的研究表明,存在一个控制核——能通过改变所有吸引子状态景观收敛到主吸引子,从而决定整个系统命运节点的最小子集。对于裂殖酵母细胞周期网络,研究表明整个网络中有4个蛋白是控制核——即当 Rum1、 we1、 Ste9 和 Cdc25 的状态固定为1、1、1、0 ,与它们所在生物功能主吸引子中的状态相同时,从每种可能网络配置开始的所有时间进化轨迹都会趋于生物功能吸引子。
信息传递与控制核的关系——以裂变酵母细胞周期网络为例
在最近的工作中(Kim et al. 2015),我们通过计算网络中每一对节点之间的传递熵来量化裂变细胞周期网络的信息架构,从结果中识别出生物学的特有的模式,并通过控制核证明它与控制机制之间的密切关系。为了区分生物系统表征的信息模式,我们将传递熵的分析结果与对随机网络集成进行的相同分析进行了比较,这些网络被构建为与每个酵母网络共享某些相同拓扑特征,但不执行相同的生物功能。
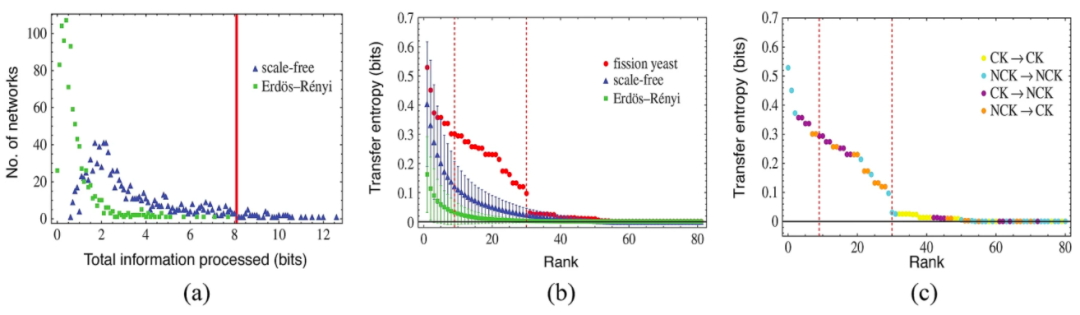
为了进行比较,我们利用了 Erdös-Rényi (ER)和无标度(SF)两种类型的随机网络集成。ER随机网络与裂变酵母网络,除了网络大小外没有共同的结构偏向。相比之下,SF随机网络集成与细胞周期网络一样,每个节点保持相同数量的激活和抑制连接,这使得SF与生物网络有相同的拓扑度分布。我们将单个网络处理的总信息定义为其中所有节点对之间传递熵的总和。结果显示,酵母细胞周期网络处理的信息量比任何集合中大多数随机网络都多(图6a)。这是一个直接测量结果,支持了与信息处理和临界点相关的假设,即在生物系统中临界点附近存在着信息传输的优化,不像以前没有基于经验数据集的定性或一般模型的研究,在这里这点被定量地清楚展示了出来。
通过根据每对节点传递熵的测量值分析缩放标度,我们还研究了处理后的信息在各个网络中的分布。标度关系表明,与随机网络相比,生物网络具有明显异常的值(图6b),生物管理信息最显著的特点是,生物网络中信息被处理得更加均匀。标度模式还表现出了统计学上独特的高-中等级节点对范围,与随机网络中相应尺度的节点相比,生物网络的传递熵明显更大。并且我们发现,这种生物学上独特的范围是由控制核节点和非控制核节点之间的信息处理主导的(图6c)。
细胞周期网络在信息结构上表现出与随机网络相区别的特征模式,控制核节点通过控制有关结构的全局因果(状态转换图中的因果流),在网络信息传递中起主导作用,调节着系统全局状态转换。
图5:裂殖酵母细胞周期调节的布尔网络模型。节点代表调控蛋白,边缘表示节点之间的两种生化作用:激活(以箭头结尾)和抑制(以条形结尾)。其中红色节点是控制核,当固定在特定值时,它可以调节网络的全局行为。该图根据 Davidich 和 Bornholdt(2008)和 Kim 等人(2013)的数据重新生成。
图6:裂变酵母细胞周期网络中的信息架构和因果结构。a ER(绿色)和 SF(蓝色)网络集成处理总信息的分布。每个数据点表示在x轴上,传输的给定信息总量的各个集合中的单个网络的数量。b 裂变酵母细胞周期中成对节点间传递熵的标度,ER和SF网络分别用红色、绿色和蓝色表示。对于1000个网络的样本,计算每个随机网络集合的平均值和标准差。虚线之间的区域表示裂变酵母的信息标度在生物学上独特的范围。c 裂变酵母细胞周期网络信息传递的标度关系。CK 表示控制核节点,而 NCK 则是一组不包含在控制核内的节点。所示数据与中间面板中裂变酵母传递熵的标度模式相同。根据一对中每个节点是在CK还是在NCK中,标度模式被分为四类信息传输。这些数字来自 Kim 等(2015)(在线彩色图)
到目前为止,我们主要讨论的都是生物学的例子,但为了进一步实现理解生物集群独有特征的目标,我们还必须理解所关心这些特性在非生命系统中出现在哪里。例如,有些非生命的、纯物理系统允许我们在没有生物组织的情况下研究可控性。引力体(Gravitating bodies)、范德瓦尔斯气体(Van der Waals gases)和自旋玻璃(spin glasses)都是潜在作为纯物理复杂系统的例子。不幸的是,这些系统中的绝大多数从未进行过任何形式的可控性分析,因为几乎没有控制它们的实际需求。耦合量子系统是个例外,由于量子计算的潜力人们对其可控性进行了严格的核查。
尽管对量子系统可控性的全面回顾超出了本文的范围,但值得指出一些与网络可控性相关比较突出的结果。最值得注意的是,许多耦合的量子系统只需通过操纵一个或两个量子比特就可以实现完全可控性,其作用类似于网络控制理论中的驱动节点。若通过使用某种形式的外部控制可以访问整个希尔伯特空间,那么一个量子系统就被称为是完全可控的(Schirmer et al. 2003)。
给定足够的外部控制量,任何系统都应该是完全可控的。例如如果可以控制一个系统每个单独的自旋,那么一个耦合的自旋-1/2粒子链将是在极其精细下完全可控的。更有趣是,为了完全控制系统,量子系统需要的最小外部控制是什么,这与Liu 等人(2011)等人在网络可控性方面提出的问题一样,答案也惊人的相似。具体来说,Burgarth 等人(2009)研究了一种通过各向同性海森堡型(isotropic Heisenberg-type)相互作用耦合在一起自旋为-1/2的粒子链的完全可控性条件,发现通过两个非对易(non-commuting)的外部控制对链末端单个节点的状态,就足以对网络实现完全可控性。
类似地,在具有XY型相互作用的海森堡型自旋链中(Z自旋分量之间没有耦合),只需操纵单个位点及与邻近位点的相互作用就可以实现完全可控性(Schirmer et al. 2008;Kay & Pemberton-Ross 2010)。然而也并非所有自旋链系统都如此容易控制。例如,伊辛型(Ising-type)的相互作用需要对每个局部自旋进行控制才能实现完全可控(Wang et al. 2016)。事实上,就像网络可控性一样,很难判断给定的拓扑结构和交互类型是否能在少数驱动节点上产生完全可控性,量子可控性领域的许多研究都集中在解决这个问题上。
现在还很难知道量子系统的非经典性是否会影响我们对纯物理系统控制的理解。但可以明确的是,这些系统中的一些系统要么通过少量的局部相互作用实现全局控制,要么需要对每个组件进行操纵。这与”控制核”部分提到的对复杂网络可控性的研究是一致的:与随机网络相比,包括生物网络在内的无标度网络往往需要更多的驱动节点来实现完全可控性,但又少于完全断开连接的网络,仅仅需要对每个节点进行操纵。这可能表明,生物系统的可控性特征程度在于临界状态,与很多时候生物系统动力学中的临界的状态相似(见“临界性”部分)。因此总的来说,在对生命群体进行分析的同时,对物理集群系统进行更广泛的分析,可能会被证明更富有成效,可以准确确定哪些特征是生命中物理所特有的,而非作为物理宇宙中一个更广泛的属性。
我们当前对所有尺度生命更深的理解,都正趋向于将决定生命属性的概念定义在信息上(Nurse 2008),即使它可能并非真的最重要。正如本文已经部分回顾那样,信息理论的工具已经被用来解释各种生物现象,从大脑神经元的放电(Honey et al. 2007;Vakorin et al. 2009;Ito et al. 2011;Lizier et al. 2011),到鱼群的行为(Butail et al. 2014,2016;Mwaffo et al 2017;Crosato et al. 2018),再到细胞内的化学信号传递(Chong et al. 2011;Rhee et al. 2012;Selimkhanov et al. 2014)。与此同时,要实现真正的“生命系统物理学”,需要对生命有比目前已知的更基本的理解(Bialek 2012)。其中悬而未决最具挑战性的问题之一是缺乏区分生命系统和非生命系统的可量化指标(Clel& & Chyba 2002; Davies & Walker 2016)。虽然现代生物学的大部分研究内容,可以在没有对生命是什么(以及不是什么)有深入的数学理解的情况下进行,但在某些科学领域,客观的、可测量的生命构成标准是绝对关键的:这些标准包括生命的起源和对外星生命的探索(Walker 2017)。在我们努力在实验室(Cronin & Walker,2016)或其他星球(Walker et al. 2018)寻找新生命实例的过程中,可量化的指标将成为决定性因素——允许在实验室中设计可进化的化学系统,穿越从非生命到生命的小径,或允许对生命进行明确探测,即使它可能存在于与已知生命完全不同的化学成分中。
在建立对生命的定量理解方面,最大的障碍之一是我们缺乏适当的控制来隔离生命过程的物理机制。虽然标准物理学对生命的运作有了一定合理的理解(Schrödinger 1992; Hoffmann 2012),但我们迄今为止还没有设计出有效的实验隔离,证明生物系统和非生物系统属性是否能够进行直接比较,或是否能解决非生命和生命系统之间集群行为的差异。亦或许没有这样的区分更有意义。为解决研究生命过程中物理机制需要更好的控制的问题,我们提出了一套想法,建立了一个框架,在这个框架中让生命存在于一个连续谱上,从我们目前认为可能表现出一些“类生命”行为的非生命系统,一直到行星规模的文明。这是生命可能存在于一个光谱中想法的自然结果,在这个光谱中,其中一些系统比另一些更有生命力,关键的区别属性是它们的信息架构。虽然我们认为这样尺度的生命可能存在,但还没能为这种尺度建立适当的度量标准,也没有在上面放置特定的例子。
其中面临的挑战之一是,对许多群体系统之间的比较分析仍有待严格制定和界定。我们认为信息论方法是最有前途的,因为它们捕捉到了物理系统在时空中存在相关的因果结构,提供了一个足够抽象和严格的数学框架,是我们发现生命过程的各种物理介质和尺度中量化生命的候选方法。而为了做出群体决策,个体必须共享信息,因此对群体系统中信息流的严格量化是可能提供新见解的一种途径,使人们了解新的计算类型(例如决策)如何从单个代理的行动中产生。
随着人工智能和信息理论应用的发展,现在可以通过许多不同的渠道提取生物行为的算法来处理生命系统中的信息,例如运动、形态学和生物电场。然而,尽管在概念和技术应用方面都取得了进展,但尚未进行系统的分析,直接将生命系统的信息特性与非生命系统进行比较,对生命系统的信息特性进行量化。大多数生物模型都是用硅片树脂或机器人建造的,这意味着不可能直接将这些模拟系统的物理性质与生物系统进行比较,毕竟后者存在于潮湿、混乱的化学环境中。
这在很大程度上限制了当前人工生命模型区分表现为“类生命”和严格作为生命物质的能力。
“一个例子:行为相似但机制不同的跨集体的信息架构”一节中的例子表明,如果它不能捕获局部级别的规则,那么不同系统中相同模拟的行为可能是不够的,因为信息流将有所不同。因此,如果只是天真地构建硅片模拟系统,可能不足以理解真实生物过程中的物理信息流。我们已经证明信息理论对特定规则选择是敏感的,而在计算机中建模者会显式地选择规则,这就使得计算机模拟生命中的信息结构与真实世界生物系统相比会直接存在差异。特别是,真实生命不允许孤立系统中的信息流结构(译注:如封闭的计算机中),不会超越或超出物理和化学法则进行外显编码(译注:如人工智能软件系统的特殊规则),我们假设这是理解生命物理学的关键一步(Walker&Davies 2013)。
此外,计算机模型通常是真实世界过程的低维表示,因此不能捕捉到生物体们处理信息的所有多重信道。向前推进的一条可能道路是建立活体系统的物理模拟。例如,油滴混合物中演化的集体行为提供了一种非生命系统物理中模拟生物特性的方法,而不仅仅是在计算机中模拟(Taylor et al. 2017;Points et al. 2018)。因此,解决我们在这里提出的问题的一条途径是开发活体系统的物理模拟,作为探索信息流物理的一种手段。
当然,要确定这种方法是否会最终能取得成果,还有很多工作要做;但我们认为,为理解集群行为,何时可归因于生命过程、何时又不能,朝这个方向迈出步伐的努力,尤其是研究并提供新的定量理论和工具,将源源不断提供新的见解并促使生物学持续发展。
https://link.springer.com/article/10.1007/s12064-020-00331-5
集智斑图顶刊论文速递栏目上线以来,持续收录来自Nature、Science等顶刊的最新论文,追踪复杂系统、网络科学、计算社会科学等领域的前沿进展。现在正式推出订阅功能,每周通过微信服务号「集智斑图」推送论文信息。扫描下方二维码即可一键订阅:

点击“阅读原文”,追踪复杂科学顶刊论文