本篇推文来自集智俱乐部的网站集智斑图的一篇路径《神经元模型:从离子通道到计算》,路径基于神经动力学模型读书会第一季臧蕴亮老师的分享整理而成。感兴趣的朋友可以结合本次分享的回放(回放链接https://campus.swarma.org/course/4433)进行学习。
分享内容简介:大脑中信息传递主要是通过底层的神经元产生并传递不同形式的脉冲来实现的。因此了解神经元如何处理、传递信息对理解环路以及脑实现相应功能的机理有着重要意义。主讲人从常用的神经元建模方法及其缺陷讲起,然后介绍不同种类神经元的计算特性(如type I和type II),并围绕这些特性以及相关论文,重点讨论基于神经元模型的研究方向及进展,最后对神经元模型的研究热点进行展望。
学习难度:难度适中
阅读基础:简单了解神经元电生理、神经元树突计算、神经网络大规模环路仿真的知识会对理解大有裨益。
臧蕴亮 | 讲者
Erfan | 整理
邓一雪 | 编辑
神经元的结构特性与脉冲放电过程
Hodgkin-Huxley model
Compartment model
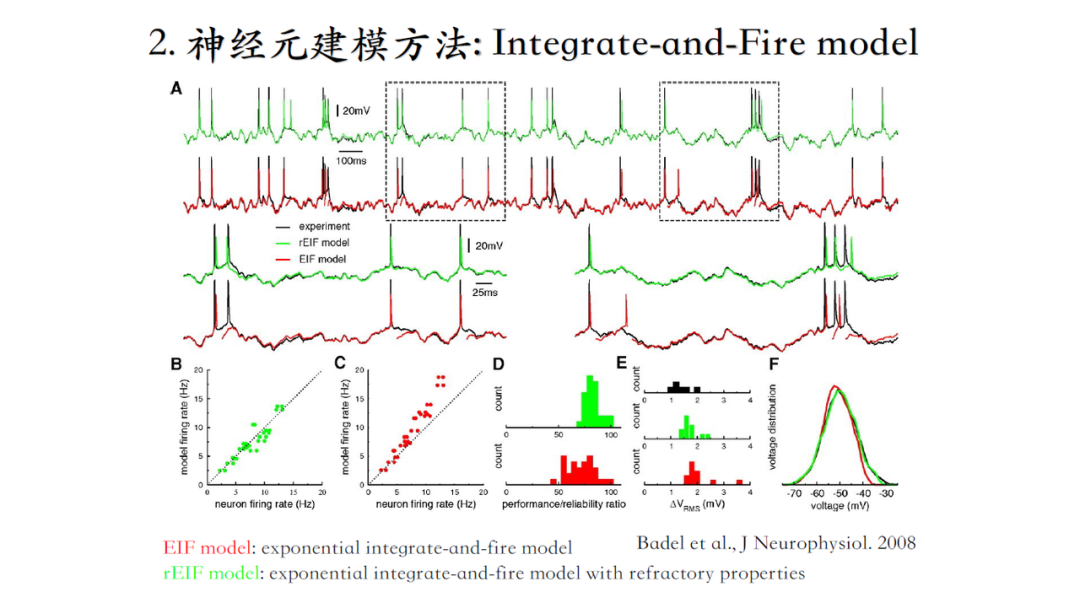
Integrate-and-Fire model
如何选择模型
与适时状态相关误差学习
神经元多路复用编码
神经元功能集群动态形成
个体差异性与稳态可塑性
神经调控不同方向
稳态可塑性之结构变换
模型简化到大环路
更微观的实验数据
物种的区别
更复杂的计算功能
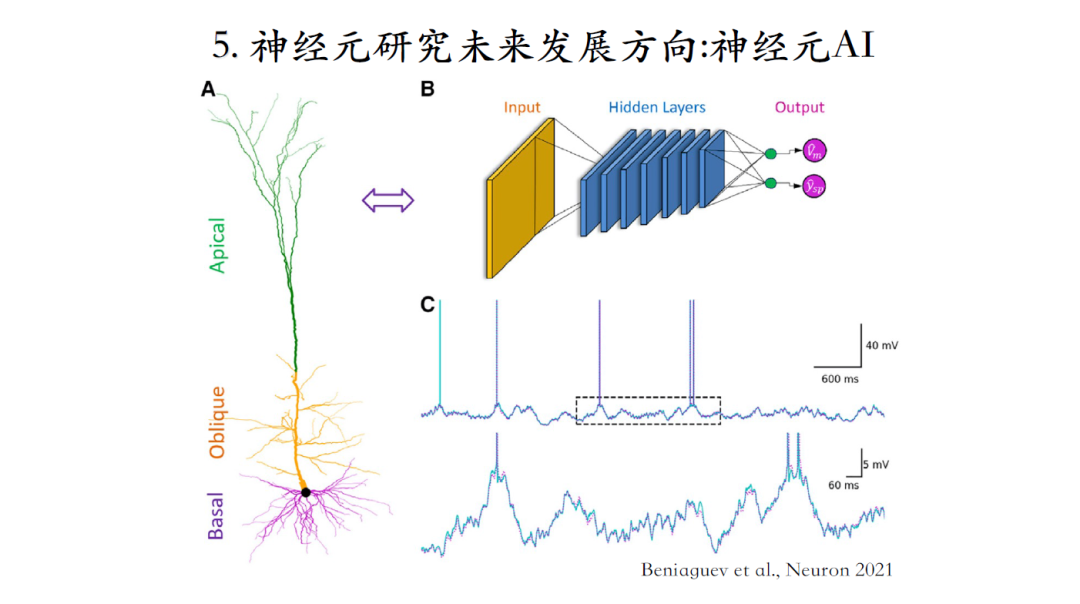
神经元AI
传统理论认为,大脑具有不同的分区,各司其职,因此对脑的研究一般以脑区为单位。但近些年发现脑区的功能划分并不绝对,远比现有认知更复杂。比如小脑不仅负责运动相关功能,也可以参与社交、感情、决策等过程。
脑及神经元之间的连接异常复杂,比如要了解视觉实现的过程,在小鼠中半径845微米的一小块初级视觉皮质V1区就包含23万个神经元,想完全解构这些神经元的电生理活动几乎不可能,在此背景下,计算神经科学就成为了解脑工作机制的重要工具[1]。
人脑有约1000亿个神经元,神经元之间的信息传递—电或化学活动错综复杂,信息整合发生在轴突初段(Axon Initial Segment,AIS),想要了解更多可以参考关于[脑](https://zh.wikipedia.org/wiki/%25E8%2584%2591) 的百科。
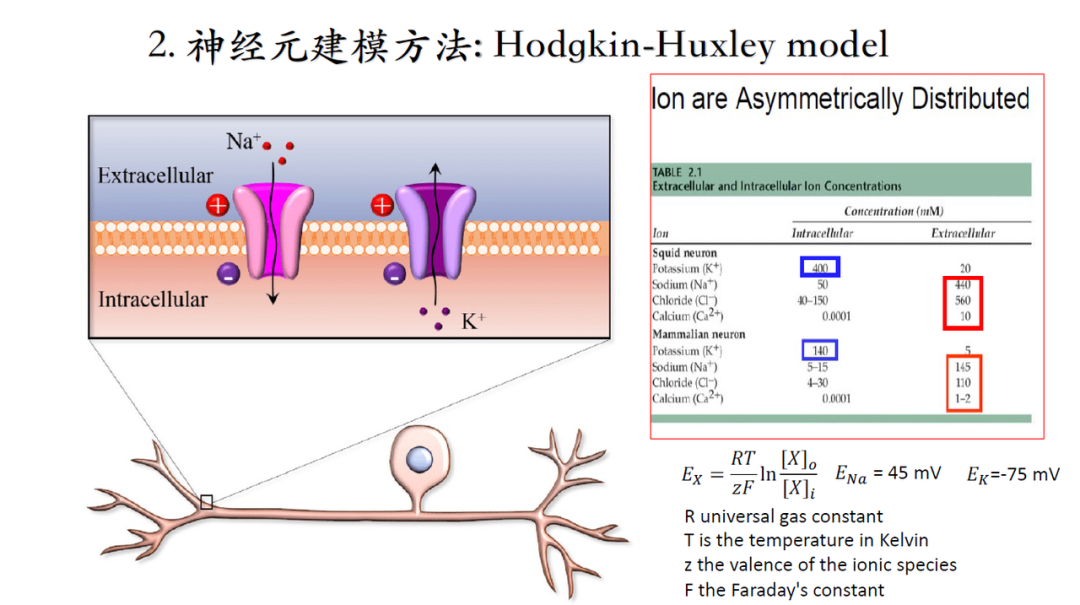
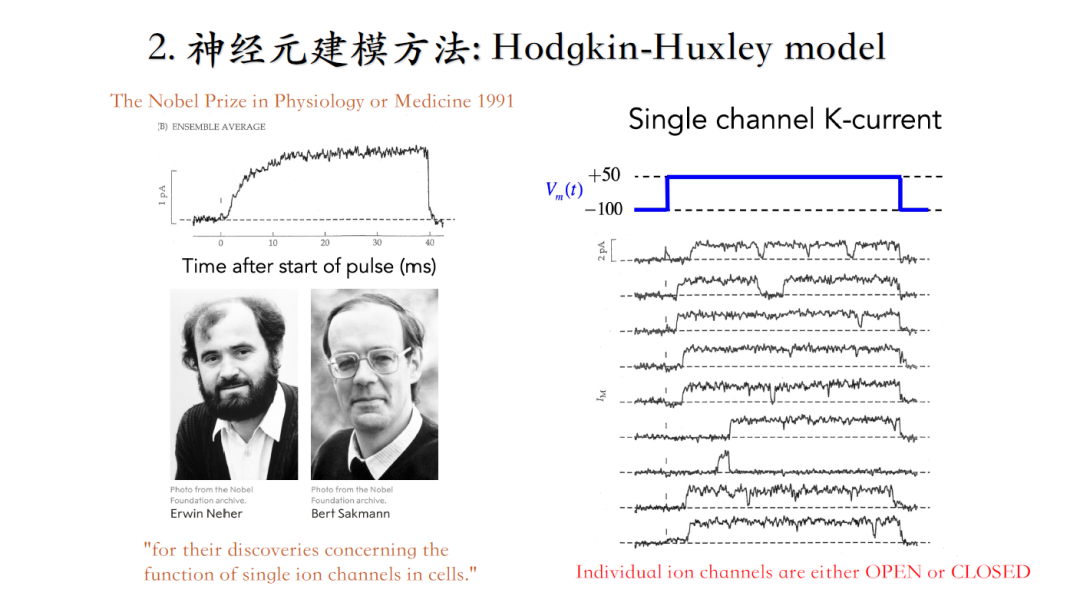
上世纪50年代,技术手段落后,大家只能在海洋动物粗大的轴突上做电信号记录,下边这篇是综合之前一系列实验测量之后关于电信号建模的集大成者,推荐大家阅读[2]。
电容器:细胞内外离子均导电,中间穿插磷脂双分子层是绝缘体,因此等效于电容器。实验中电导无法直接测量,通过基尔霍夫定律可以计算电导。
电压门控电流:细胞内电势的升高,改变了了电压门控离子通道的构象,使通道呈开放状态,允许离子在浓度差的驱动下流动产生电流。
电势差:细胞内外多种离子浓度的差异(由于细胞膜的分隔)导致电势差的出现。通道打开,势能可以转化为电能。
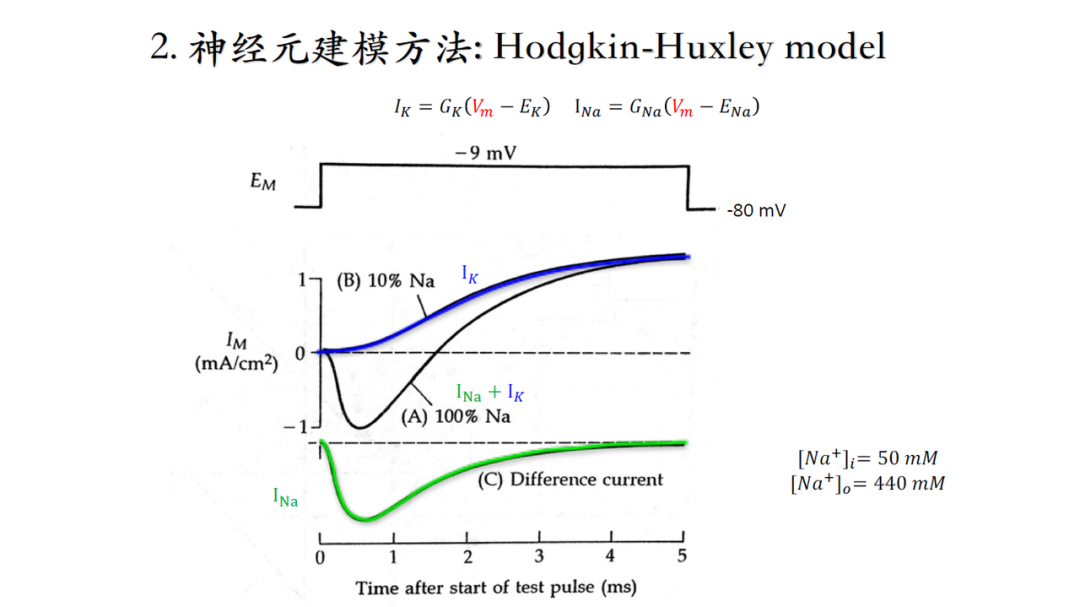
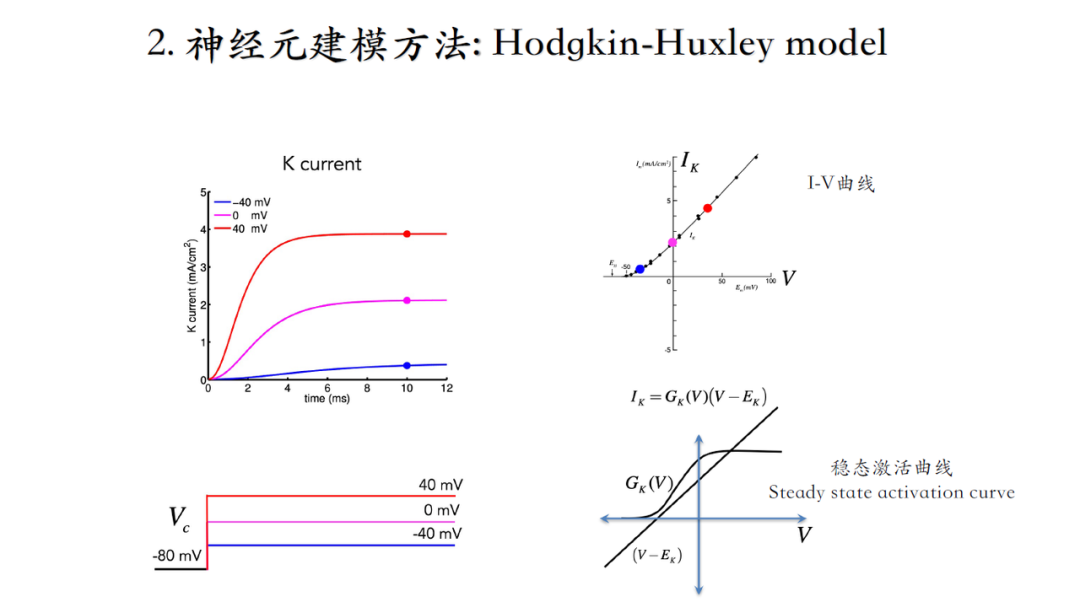
分离钠电流以及钾电流(当细胞内外钠离子浓度相同时,钠电流为0,因此测得电流实际为钾电流。然后通过生理状态下测得的钠钾综合电流减去前面测得的钾电流即为钠电流),再根据电流、电压计算电导值(电流的稳态值),如下图所示。
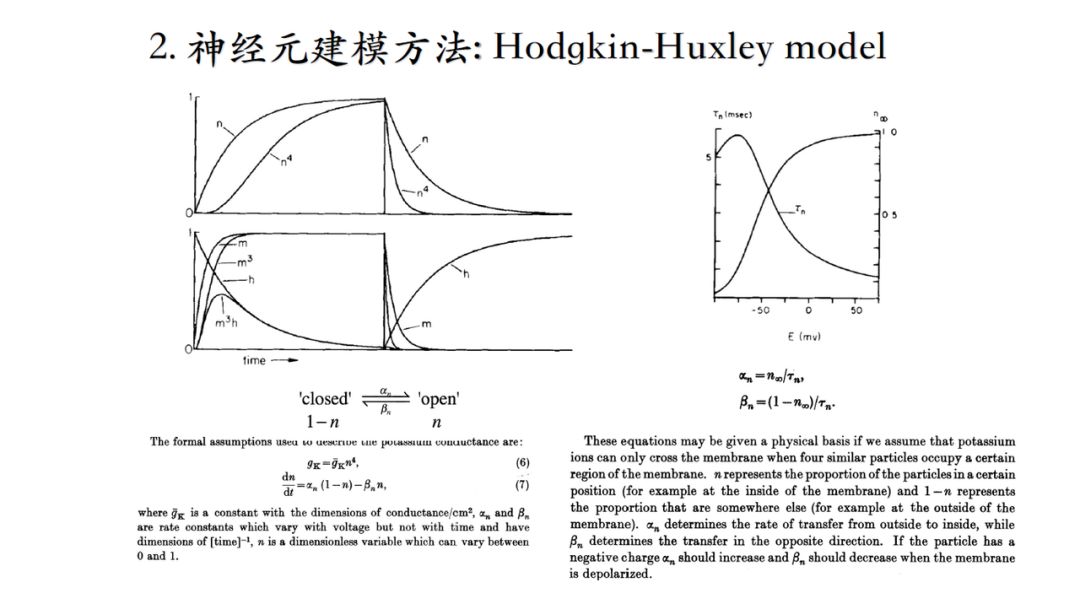
电流幅度的变化不是突然二进制阶跃变化(从0到1),而是随着时间逐渐达到稳态的,因此要对这个时间过程进行建模(当时人们并不知道单个离子通道的存在,只能从宏观上理解电流,并且将宏观电流理解成不同的particle处于0或1态,这一猜测预示着离子通道结构的存在,后续通过膜片钳技术得以证实。)。
1991年的诺奖研究,证明单个离子通道的存在,使用到了膜片钳技术。
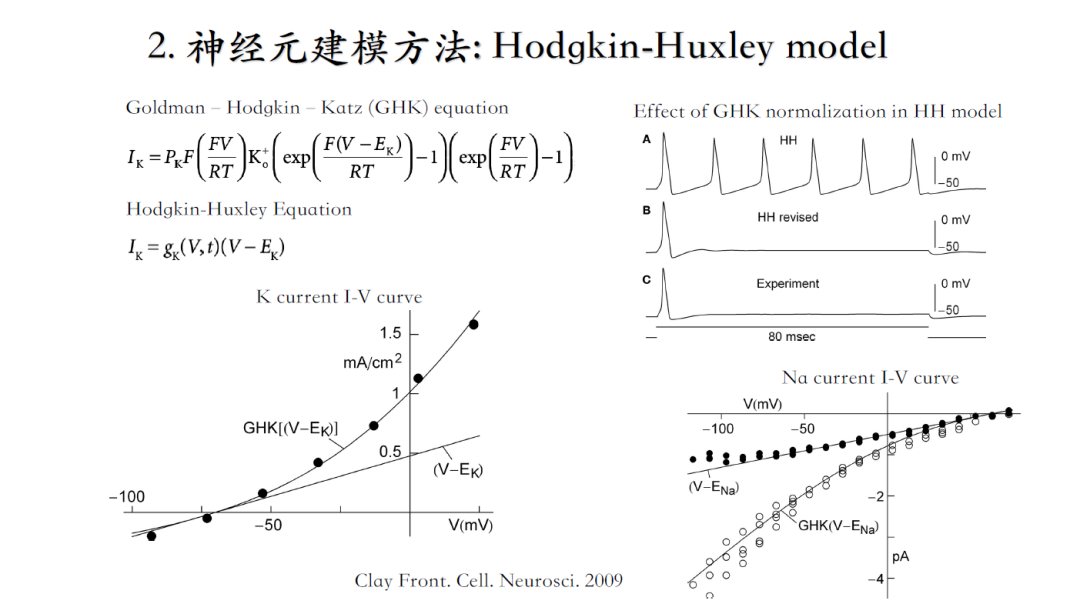
HH模型直到今天依然是我们研究神经电生理的基石,但是在某些条件下它存在着一定缺陷。Clay对比了HH模型与GHK模型的差别,发现理想情况下通过GHK模型获得的稳态激活曲线,更加吻合实验获得的曲线。但是如果细胞外存在大量钙离子、镁离子等2价阳离子,会影响钠离子的结合位点,从而影响IV曲线(使其呈线性)。在鱿鱼大轴突中存在大量的钙离子和镁离子,误打误撞使得钾电流曲线能够被HH模型完美拟合。而哺乳动物体内的IV曲线可能介于直线与曲线之间。因此,针对不同研究目的需要仔细考量两种模型的选择[3]。
过去的研究者(John Eccles等)认为树突仅仅接收信息,由胞体处理信息,以胞体为中心[4]。
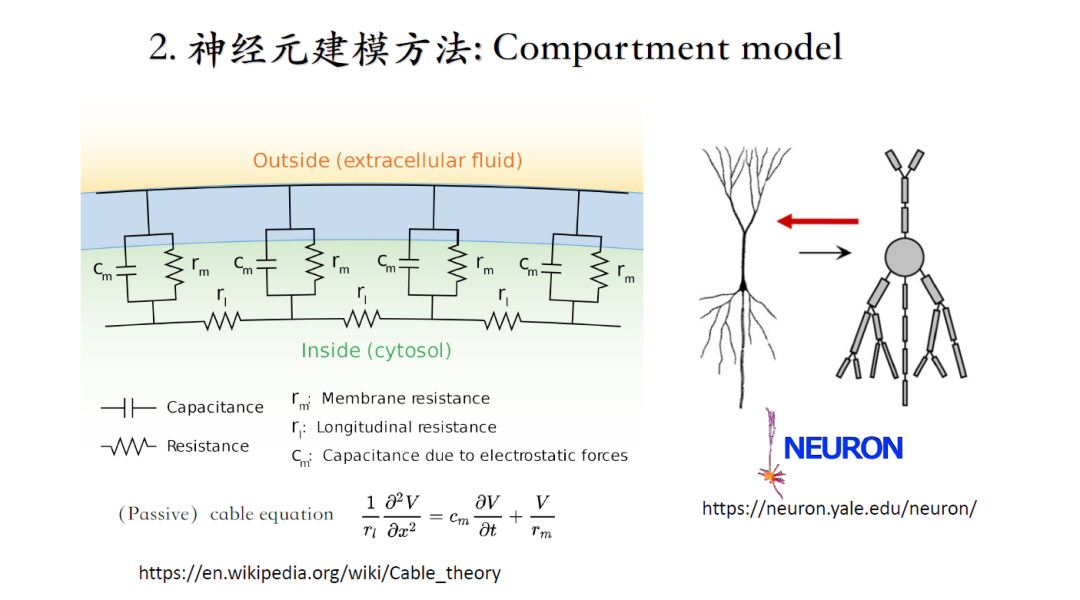
Wilfrid Rall提出了[Cable_theory](http://en.wikipedia.org/wiki/Cable_theory)(Compartment模型),将神经元的几何结构离散成许多空间小单元,最开始没有考虑离子电流,每个小单元等效为一个被动的RC等效电路。不同小单元之间通过电流的扩散过程,使电信号传播。这里可以近似成一维方向的传导,因此可以用单个变量的偏微分方程去描述。反应-扩散方程可以自己通过有限差分法求解或者通过NEURON软件调参求解。
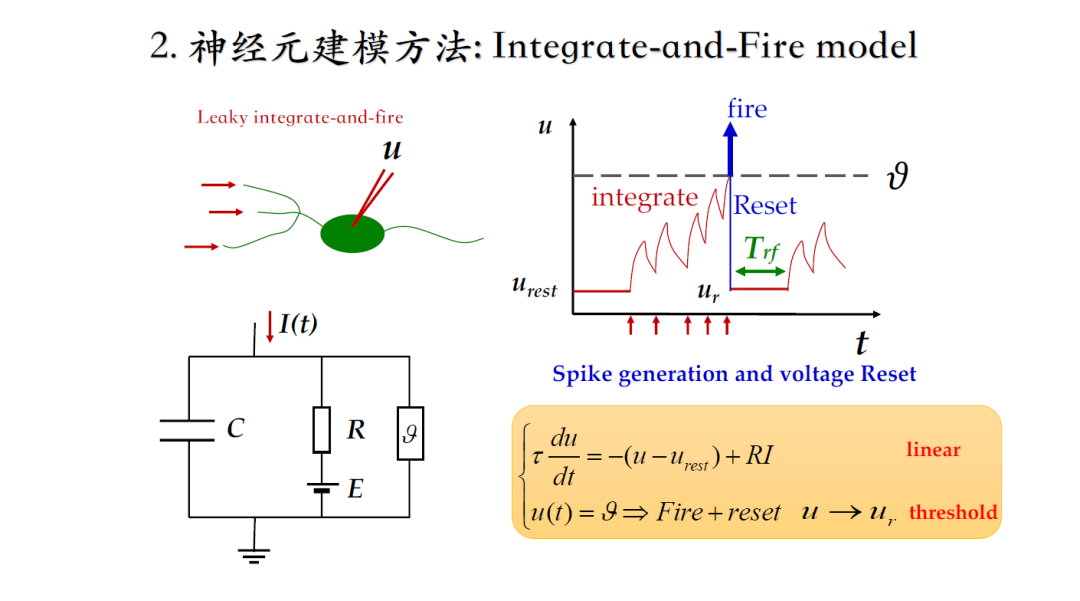
整合发放模型,广泛应用在大规模环路仿真和脉冲神经网络中。
但是它没有离子电流,神经元接受足够强足够密集的刺激后,有一定几率使电势差达到脉冲激发阈值,进而激发一次脉冲,接着回到静息水平。
下边这篇文献没有任何离子通道,仅关注神经元接收什么信息,是如何连接的,但是通过改进整合发放模型,可以使脉冲心态与实验数据非常接近[5]。
从左到右,生物现实性(接近真实神经元的生理特性)逐渐增加,计算复杂度也越来越大。选择什么模型取决于关心问题的尺度,关心哪方面特征。
最新研究表明,环路中如果引入离子电流,会使环路呈现的脉冲激发特性更接近实验观测,说明离子电流对于大脑工作机制十分重要,https://www.youtube.com/watch?v=O1_0KzVTEgE。
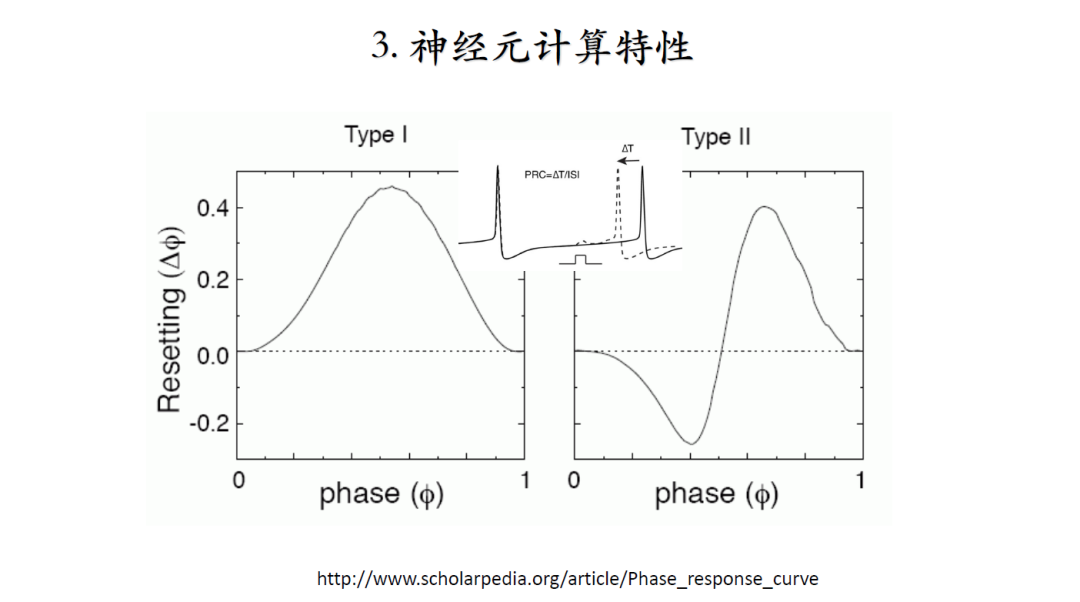
神经元的分类无唯一标准,例如可分为兴奋性、抑制性,静息态和自发激发态等。根据神经元发放脉冲后,脉冲频率和电流曲线是否连续可将神经元分为Type I和Type II。如果曲线连续则为Type I,不连续则为Type II。对于Type II, 当电压低于脉冲阈值时,会呈现出共振/振荡效果,由于Type I的连续特性,这类神经元可以有效地编码输入信号的强度[6]。
[相位反应曲线](http://www.scholarpedia.org/article/Phase_response_curve)的定义:在能够产生规则脉冲的神经元中,于两个相邻脉冲间不同时间点施加微弱的刺激,下一个脉冲的时间会被如何改变,提前还是延后,总是提前则为Type I。如果刺激位于相位晚期脉冲被提前,于早期脉冲被延迟,这类神经元则为Type II神经元。
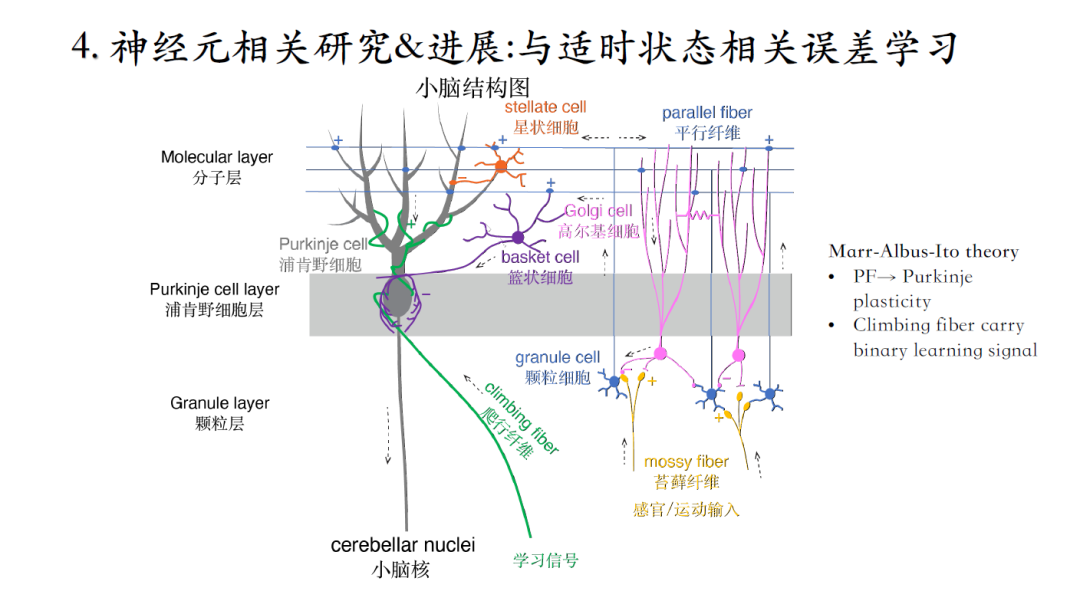
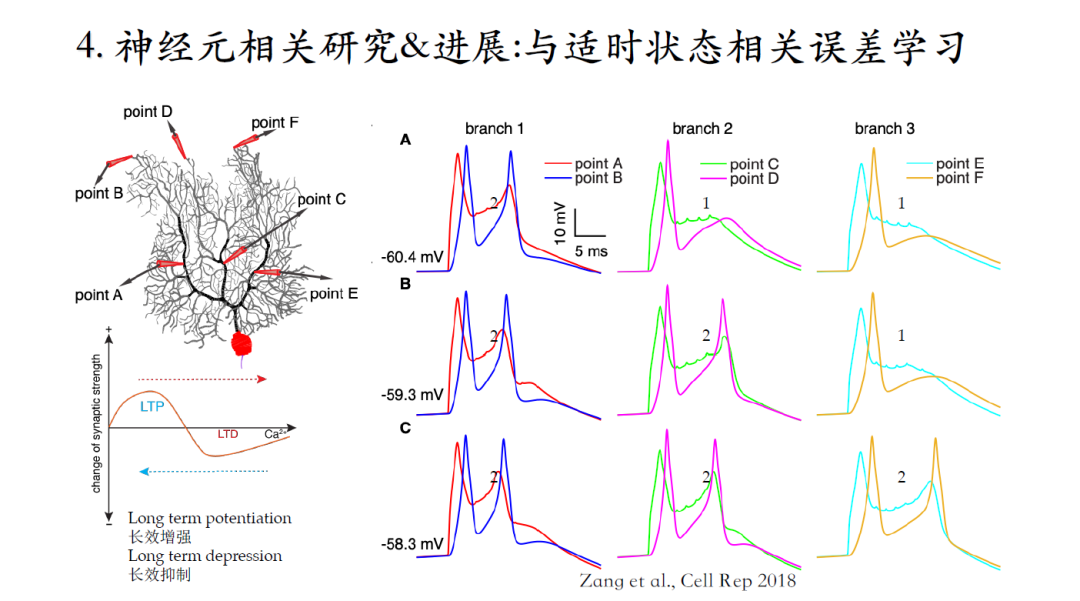
小脑中的浦肯野细胞是小脑皮质的唯一输出,运动感官信息经过苔藓纤维传入,再经过平行纤维传递给浦肯野细胞的树突,小脑输出的运动状态信息与大脑预期不一致时,通过爬行纤维传入误差学习信号,进而改变平行纤维与浦肯野细胞的突触连接强度来实现小脑学习。
过去大家认为爬行纤维给浦肯野细胞提供二进制的误差学习信号,这种假设似乎表明小脑学习效率的低下[7-8]。
由于实验技术的限制,通过实验手段很难系统研究这个问题。为了解决这一问题,需要能整合大量实验数据的新模型。计算生物学领域常见的方法是在大量细胞中收集不同离子电流特性的数据,进行平均处理,然后再用这些拥有均值特性的离子电流构建模型,去模拟平均化之后的模型输出信号。但是这种平均处理的方法从原理上是错误的,并不能总是获得满意的模型,以浦肯野细胞为例,之前十余年Erik De Schutter小组一直无法得到满意的模型。臧老师自己的模型放松了每个参数的约束,虽然采用了平均处理的理念,但只要参数值在测量的范围之内就可以,这样就得到了基于Compartment的新浦肯野细胞模型[9]。
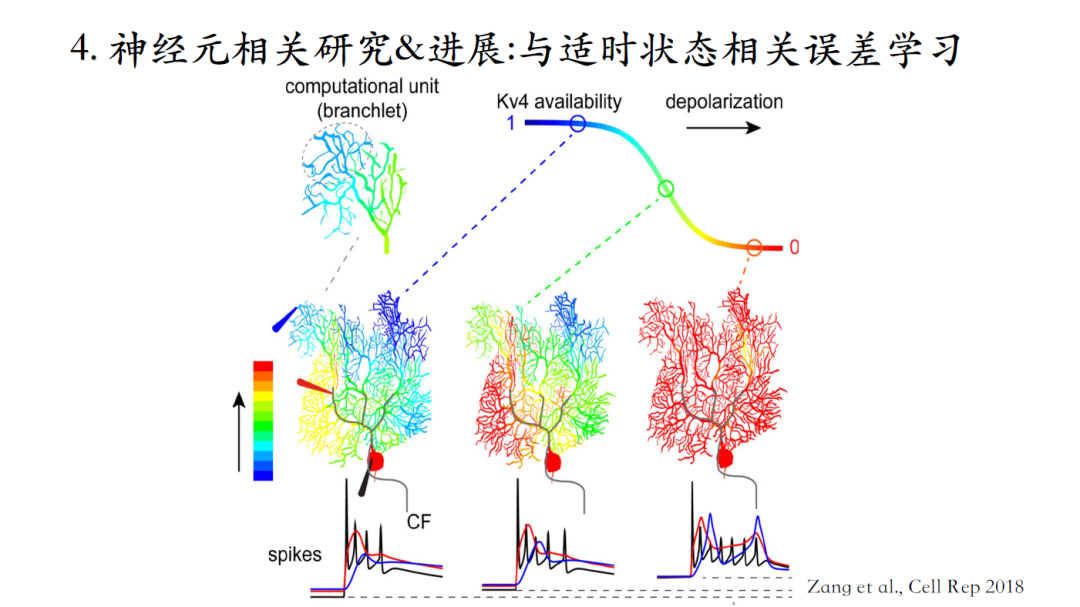
爬行纤维在浦肯野细胞中的树突响应与浦肯野细胞的兴奋状态相关。以浦肯野细胞的树突响应为例,在电势水平较低的时候,这个兴奋仅仅局限在树突主干以及离胞体比较近的区域。随着兴奋状态的提高,树突响应逐渐扩散到整个树突,这个过程的机制解释如下:
在树突的远端分布着大量的KV4离子通道,电势水平比较低的时候钾电流的availability比较大,这个电流相当于一个刹车片,阻止电信号向树突末梢进行传递。随着电势水平逐渐升高,钾电流逐渐失活,从而允许爬行纤维在树突近端激发的电活动传递到树突远端。除了树突响应,胞体输出(黑色曲线)与浦肯野细胞的实时兴奋状态也是动态相关的。
浦肯野细胞的不同树突分支也有着不同的兴奋特性。浦肯野细胞树突分支上脉冲数量有另外一层意义,代表着该位置输入的钙离子信号的浓度变化幅度。如果有两个树突脉冲,就意味着有更大的钙离子浓度信号。钙离子信号是平行纤维与浦肯野细胞间突触可塑性的引发因素,钙离子信号浓度较高,则平行纤维与浦肯野细胞之间的树突突触会实现长效抑制LTD;反之若浓度较低,则实现长效增强LTP。
简单总结:爬行纤维在浦肯野细胞中的响应能够根据该细胞的实时兴奋状态产生一个模拟信号而不是二进制的有或无信号。
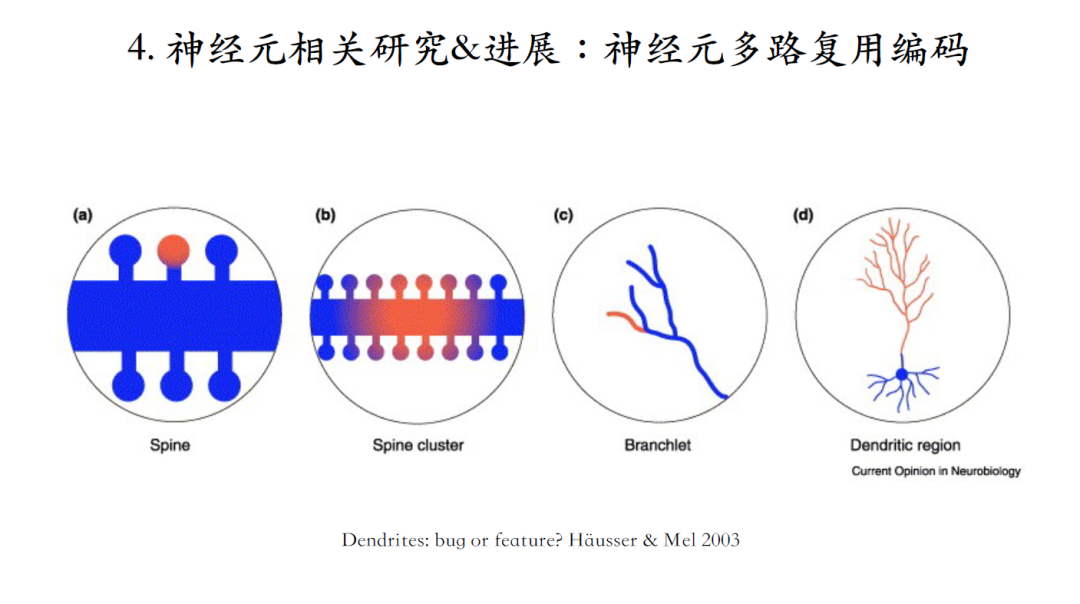
关于神经计算的基本单元,人工智能领域还是假设单个神经元是神经计算的基本单元,但是在真实的脑或者脑环路中,越来越多证据开始支持从单个树突棘Spine到树突分支在脑信息处理中的一个重要作用。浦肯野细胞是脑中已知的拥有最复杂树突结构的一类神经元,目前的理论都假定其是一个线性的点单元。下面臧老师将介绍,浦肯野细胞如何对输入的运动感官信号进行编码处理[10]。
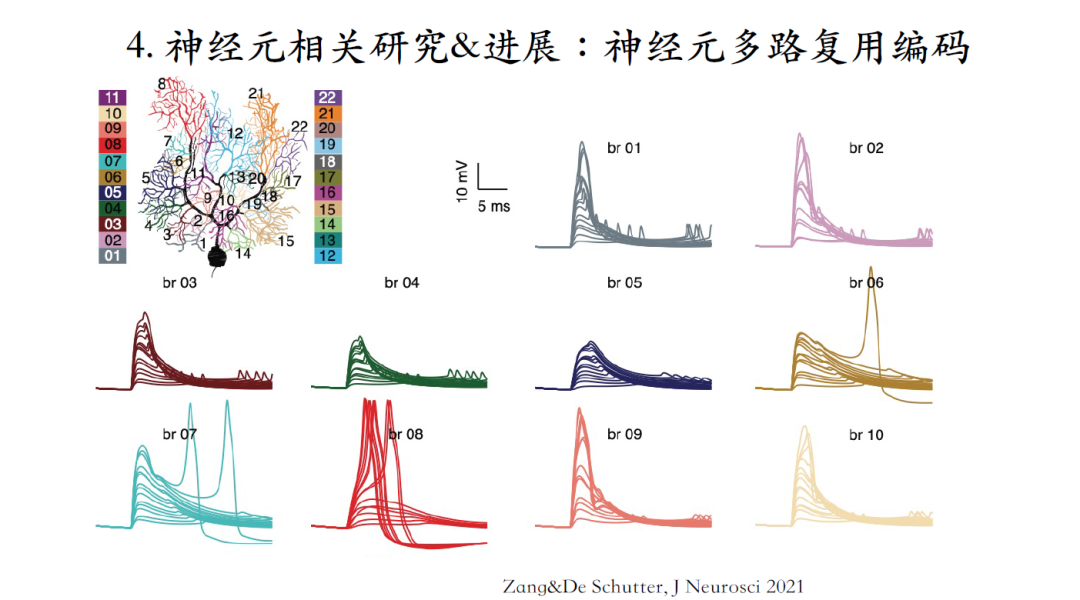
最新研究表明小鼠中平行纤维是以Cluster的方式激活对浦肯野提供兴奋信号,上图中的绿点代表模型中树突激发的突触位置。通过系统改变输入信号的强度:当信号较弱时,浦肯野细胞的树突对输入信号进行线性编码;当输入信号增强到一定幅度之后,树突响应幅度会产生一个超线性增加或者说是一个jump跃变。这意味着,在这个树突分支中产生了一个树突脉冲。当继续增加输入信号的强度,树突响应强度不再增加。而且树突脉冲仅仅局限在接收到这个输入的树突分支之内,在向胞体和其他分支传输的过程中该信号会产生急剧的衰减[11]。
根据神经元的结构特性,将浦肯野细胞的树突分支细分为22个分支,系统仿真每个树突分支的输入输出曲线特性,发现不同分支拥有非常不同的响应特性:例如树突分支8只需要激活40个平行纤维的突触就可以稳定地激发树突脉冲;而在其他树突分支如分支4、5,即使激活100个突触也无法激发树突脉冲。
将所有树突的输入输出曲线总结成一张图,发现颜色鲜艳的8/12/21/22树突分支中,输入输出曲线会呈现出非常明显的超线性增加;其他树突分支则没有明显的超线性增加。
问题:离子电流是均匀分布在浦肯野细胞的树突分支中,为什么有些树突分支可以产生超线性增加,而有些树突分支不能产生超线性增加?
为了研究这个问题,先假设每一个树突分支都能够产生树突脉冲。以树突分支8为例,用动作电位钳钳制这个树突分支与其主干连接的segment,在钳制不同的树突分支与主干连接的部位使用同一个脉冲信号,这个信号是之前仿真得到的一个树突脉冲波形。接着分析两个信号,一个是被钳制segment的轴向电流,它代表这如果这个树突分支能够产生脉冲,其余树突对这个树突分支的负载。第二个需要测量的是每个树突分支的表面积,他代表着如果这个树突分支可以产生树突脉冲,这个分支可以用来激活去产生树突脉冲的电流源。最终分析发现一般情况下能够产生明显树突脉冲的分支,会拥有较大的表面积或者拥有较小的轴向电流。但是这两个参数都不能够单独解释A图的结果,因为在其他分支中一样可以拥有较大的表面积,或者拥有较小的轴向电流。最终发现表面积和轴向电流的比例,即源和负载比例,决定了神经元不同树突分支的不同兴奋特性。
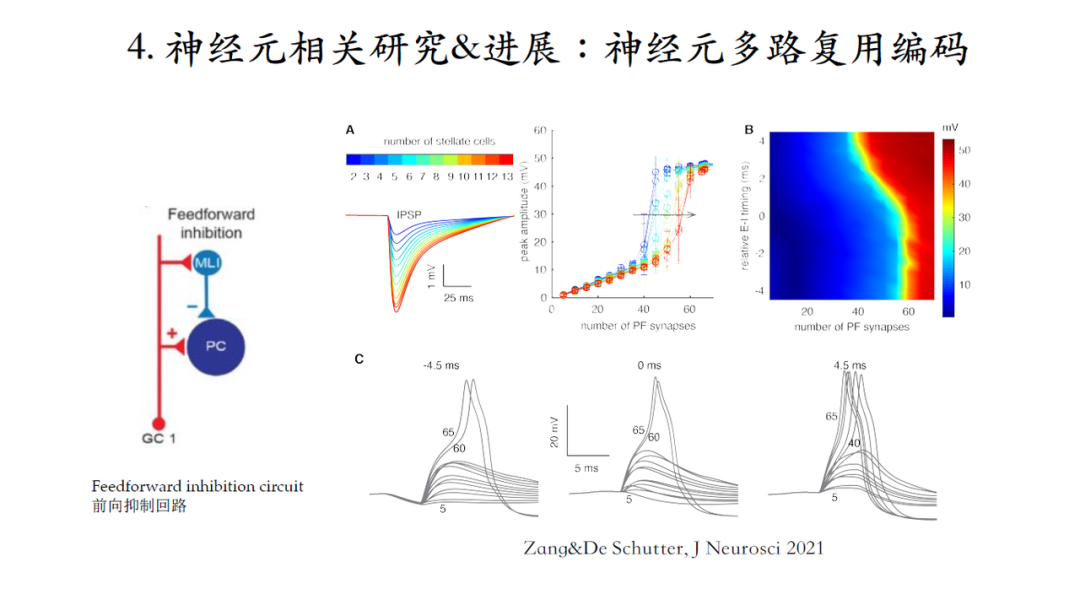
在小脑回路中,存在一个经典的前向抑制回路Feedforward Inhibition。颗粒细胞的轴突-平行纤维在激发浦肯野细胞的同时,也会激发抑制性神经元去抑制浦肯野细胞,进而延迟浦肯野细胞产生下一个脉冲。这个图研究抑制性神经元的激发如何影响浦肯野细胞对平行纤维输入信号编码。
A-B图曲线不同颜色代表着抑制强度,当抑制信号逐渐增强,树突脉冲的阈值被明显提高,相当于输入输出曲线整体右移。由此可见,抑制性神经元可以动态调整线性编码和树突编码的编码范围[12]。
前向抑制回路存在着另外一种情形,抑制性神经元是被另外一批平行纤维激活,也就是这批平行纤维并不直接与被抑制的浦肯野细胞连接。第一种模式中,抑制性输入大约发生在刺激输入的1.4毫秒之后,经典的兴奋-抑制延迟。而在第二种模式下,抑制和兴奋的相对时间则可以有不同的配置。
抑制发生时间点对抑制效果的影响:研究发现当抑制发生在兴奋刺激之前对脉冲树突的抑制性效果是最有效的。
当神经元接受来自抑制性神经元固定强度(8个)的抑制时,如果抑制发生在兴奋刺激之前的4.5毫秒,树突脉冲阈值增加到需要激活60个突触,但是抑制发生在刺激之后,抑制效果则逐渐减弱。
上述浦肯野细胞对输入输出信号的编码计算均是发生在树突局部的一个计算,下面介绍浦肯野细胞的胞体如何对平行纤维的输入信号进行处理并传递给下一级神经元。
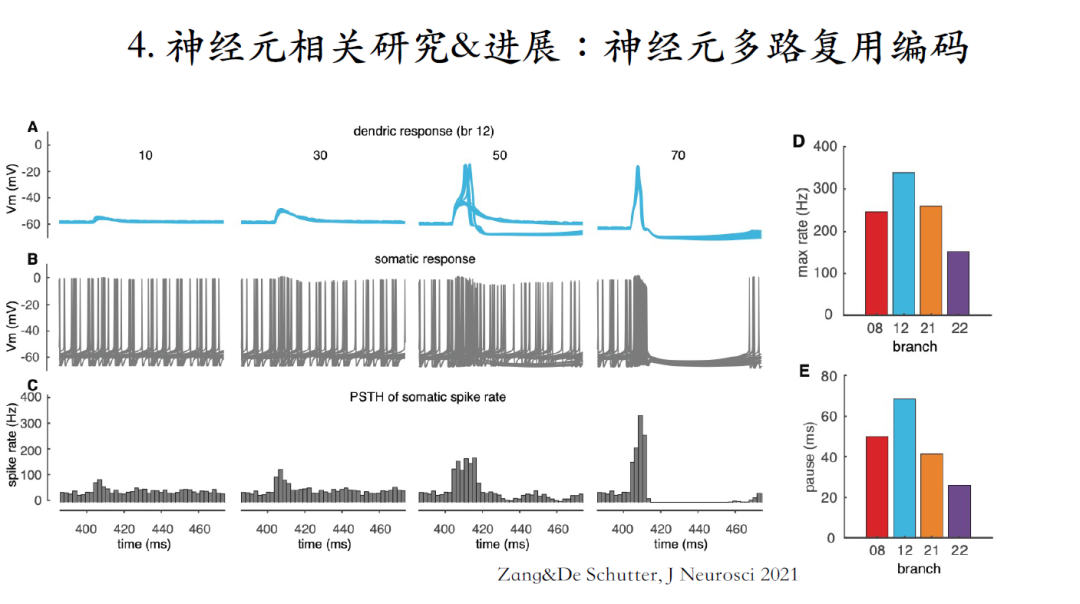
通过改变背景噪声以及突触激活位置,这里进行了大规模仿真。对每一个平行纤维输入信号的强度仿真了500次(trial),获得了一个平均脉冲输出,该协议接近于人体小脑中存在噪声情况下的生理输出。发现在输入信号较弱的情况下,树突响应信号随着输入信号的增加而逐渐增强,与之相应的胞体脉冲输出信号则会更加靠近刺激信号,增加胞体脉冲输出的瞬间频率,形成一个burst。当输入信号强度增加到一定程度之后(比如50个突触),就可以在某些Trial中激发出树突脉冲。树突脉冲激发之后,在形成burst的同时会伴随一个停顿信号,因为该现象具有随机性,并不能在每一个trial中都稳定观测到,所以在胞体脉冲平均输出频率中并不明显。当增加输入信号强度到激活70个突触后,我们能够在每一个trial里都稳定激发出树突脉冲,进而观测到稳定的停顿信号。因此神经元可以通过burst-pause信号对输入信号进行编码。
简单总结,神经元是以单个树突分支为计算单元实现神经元的多路复用代码。
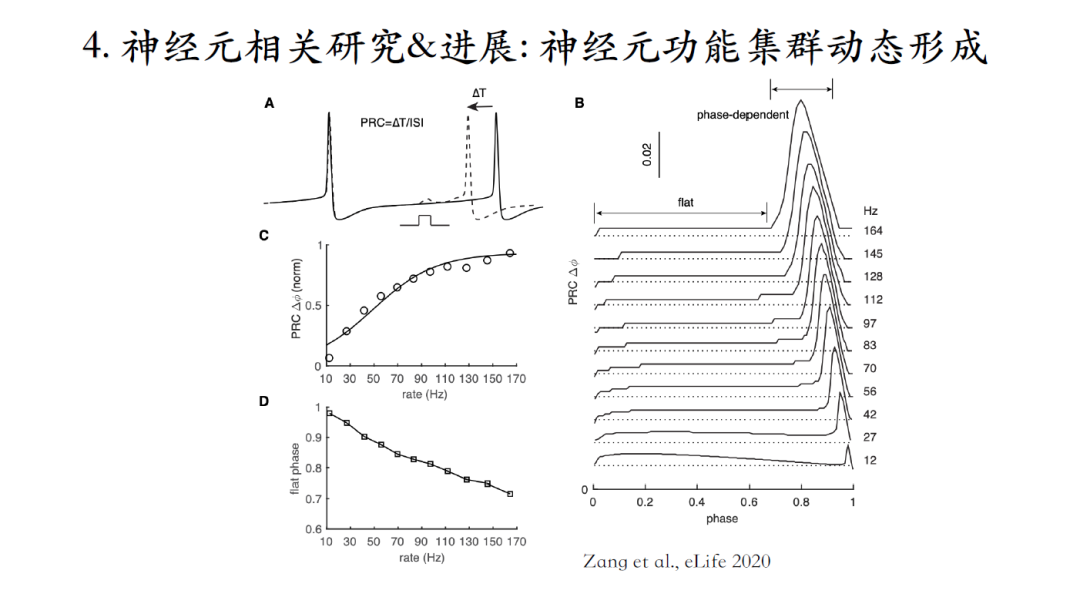
浦肯野细胞的相位反应曲线非常特别,当脉冲频率较低时相位反应曲线几乎是平的,且接近0,随着脉冲频率增加在相位末期逐渐出现一个Peak峰,当脉冲频率升高的同时,这个峰也会变高和变宽。
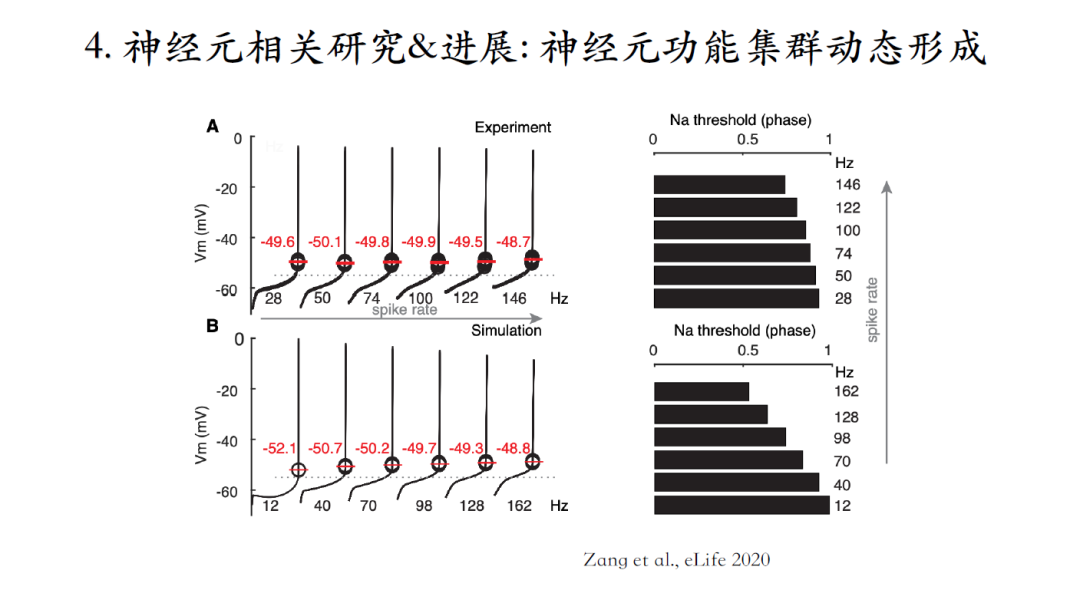
臧老师解释:在脉冲间期早期,电势水平低,离钠电流激发阈值较远,微弱的正刺激不会激发钠电流,然而在脉冲间期的后半段电势水平较高更接近钠电流激活阈值。这时给一个正刺激会激活钠电流从而加速下一个脉冲的激发,这就是在相位反应曲线末端出峰的原因。
神经元脉冲频率较高时,脉冲间期的整体电势水平比较高,会更早地到达电流的激发阈值,建模和实验结果可以互相验证。下面要探索这一发现的功能意义[13]。
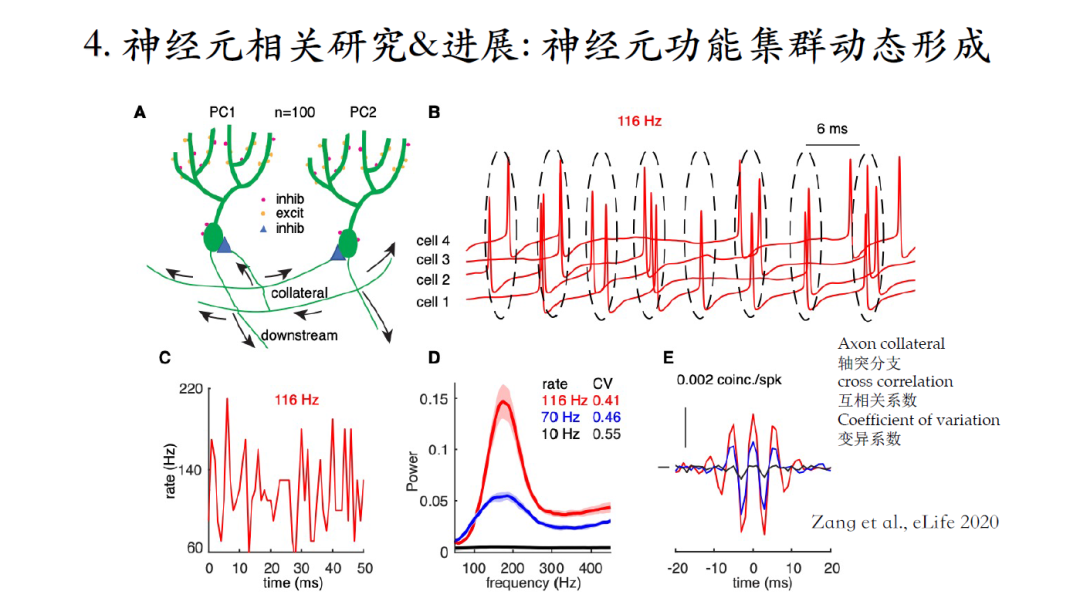
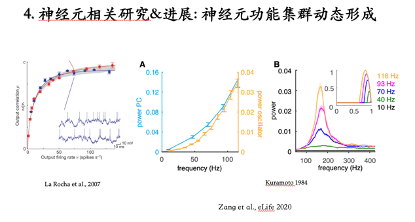
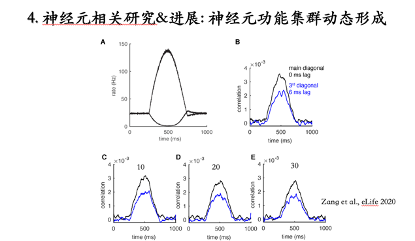
浦肯野细胞在小脑回路中能够通过轴突分支 (axon collaterals) 建立抑制性连接。根据这一现象构建了一个环路模型并研究发现:这个环路能够产生一个与单神经元脉冲频率无关的高频振荡。分析每一个神经元的脉冲激发时刻会发现不同神经元的激发时刻集中在大约间距为6毫秒的Cluster里,它不是严格意义上的Spike-to-Spike同步,而是集中在一个Cluster里的弱同步。环路振荡的频率是1000/6大约在160赫兹的水平,可是单个神经元的脉冲频率是116赫兹。所以这个环路可以产生与单个神经元脉冲频率无关的一个高频振荡,而且其震荡强度随着单个神经元脉冲频率的降低逐渐减弱。在单个神经元的脉冲频率降低到10赫兹的时候,这个网络不会产生任何高频振荡。
所有与脉冲频率相关的振动强度或脉冲同步性的变化也都可以反映在神经元间脉冲激发时刻的互相关系数中。
但是,目前的研究无法从逻辑上区分出与脉冲频率相关的高频振荡强度是由于脉冲频率变化直接导致的,还是与脉冲频率相关的相位反应曲线变化所导致。因为,前人研究(2007)表明,神经元之间脉冲输出同步性可以单纯随着神经元脉冲输出频率的增加而增强,臧老师要证明他发现的现象与上述现象是同一个机制还是另有原因[14-15]。
利用Kuramoto振荡子模型,该模型没有涉及任何脉冲频率的概念,首先证明了从因果关系上,高频振荡强度的改变与相位反应曲线变化之间存在因果联系。神经元之间脉冲输出的同步性随着脉冲频率的改变的趋势与之前2007年的结果不同。
之前的仿真结果都是在神经元达到稳态脉冲频率的情况下取得,但是这一特性如果在功能上能发挥重要的功能作用,脉冲同步性需要随着神经元的脉冲频率动态发生改变。通过模型仿真发现环路脉冲输出同步性可以实现瞬间改变,模型输出同步性会紧跟脉冲频率改变的轨迹。而且模型中存在部分异质(脉冲频率降低)神经元,也不会明显降低模型输出同步性。
总结:首次从理论上重复出与脉冲频率相关的相位反应曲线变化特性,解释了脉冲反应曲线特性的产生机制,通过环路仿真,提出了一种神经元功能集群动态形成新理论。
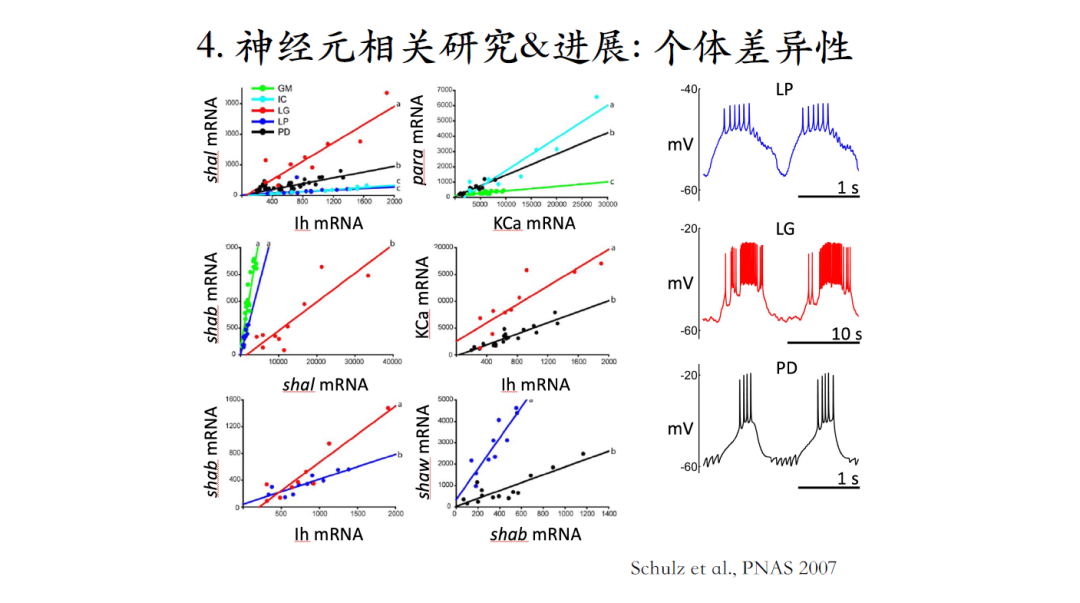
图中神经元1和2有着非常类似的激发模式,也就是动作电位特性很接近。但如果测量两个神经元中的电流会发现构成电流的幅度不同。把数据整合起来,发现同样一种电流例如钙离子电流在不同的细胞中会有3~5倍的变化,但大家容易忽视神经元的个体差异性这一客观事实[16]。
由于实验中电导测量误差较大,很多人觉得这些差异是由于测量不准确所导致。通过对表达相应离子通道的mRNA水平进行测量,最后结果发现不同细胞中的mRNA含量差异也同样很大,这也就证实了单个神经元之间确实存在个体差异性[17]。
如果用求平均的方式约束模型中的离子电流特性,多数情况下并不能获得满意的模型,即使得到了,该模型也不鲁棒(施加很小的参数改变,模型输出特性就会改变)。然而,真实的神经环路无时无刻地不在经受扰动,以离子通道为例,它作为蛋白质会经历凋亡,在神经元胞体产生的mRNA需要要运输到远端进行一个局部的蛋白质合成(有争议),不可能每次运输都那么精确。同时神经系统还面临各种环境噪声,比如体温的改变,情绪紧张引起的神经调质释放。
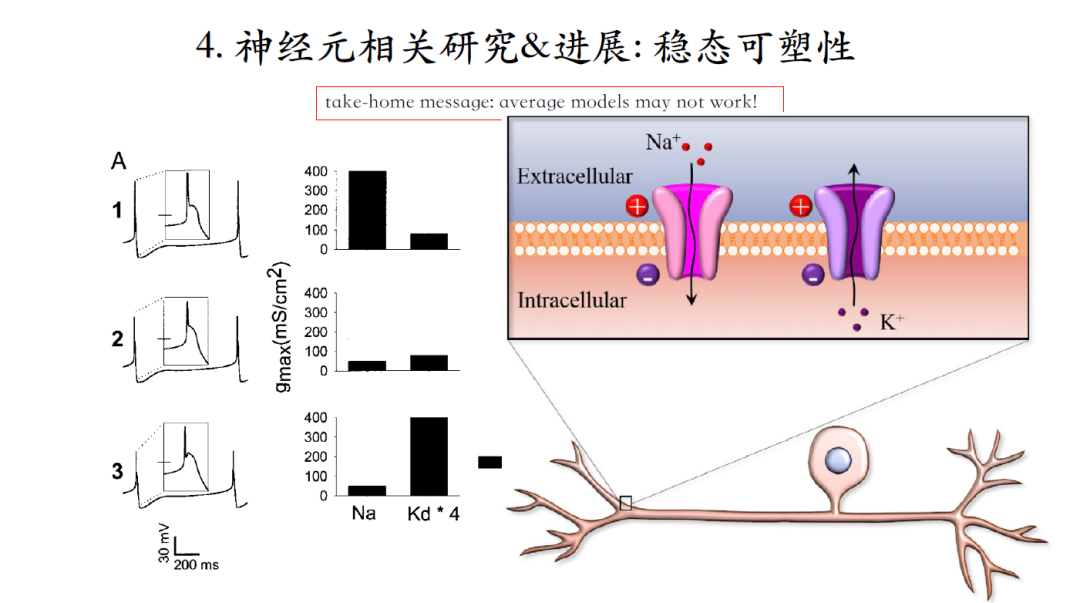
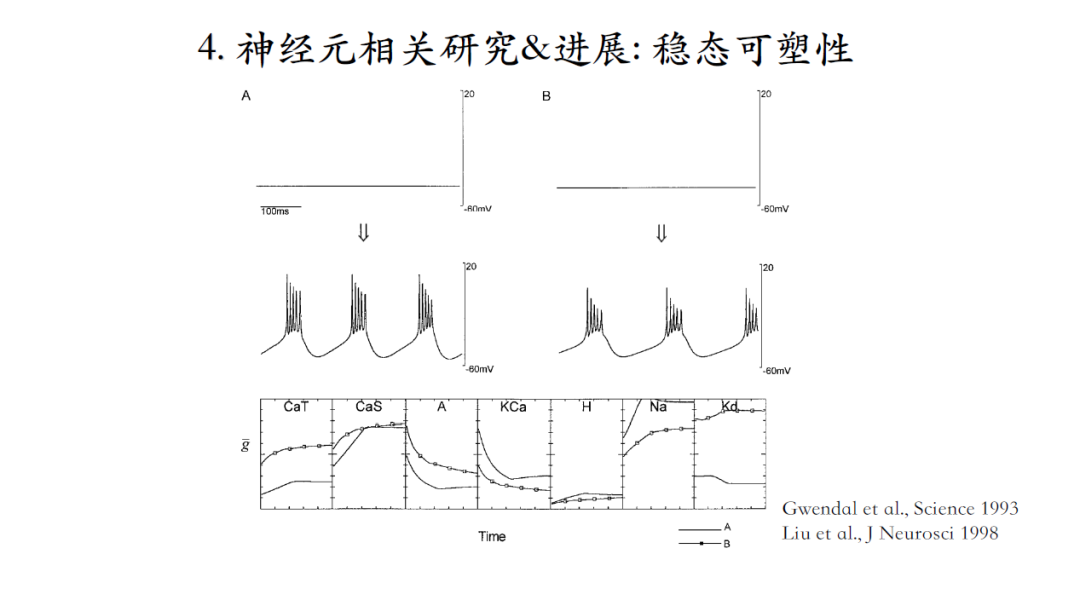
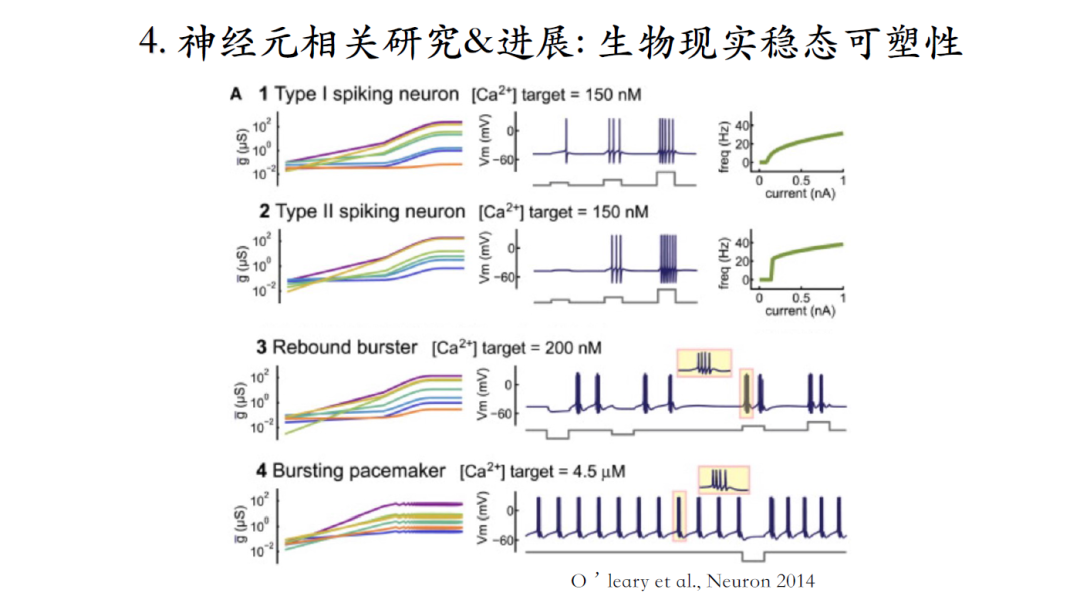
尽管同一种类神经元可以输出类似的脉冲信号,但是神经元之间存在着个体差异性,即组成个体神经元的各种离子电流组合千差万别。什么样的机理可以保证不同个体中各种电流电导值差异很大的情况下,他们组成的神经元能产生稳定的输出?
稳态可塑性是一种可能的生理机制,工作原理就是引入自动控制原理的负反馈概念,设定一个setpoint定点(代表系统想要获得的一个状态),当系统的输出偏离定点时,便会在误差的引导下,促使相应离子电流的电导值发生变化,最终使系统输出恢复到setpoint,即产生相同的脉冲输出。
图A和B两个模型初始状态对应着不同的离子电流起点幅值,经过负反馈调节之后,都可以达到相同的神经元脉冲输出模式,即便终点的离子电流幅值也依然不同[18-19]。
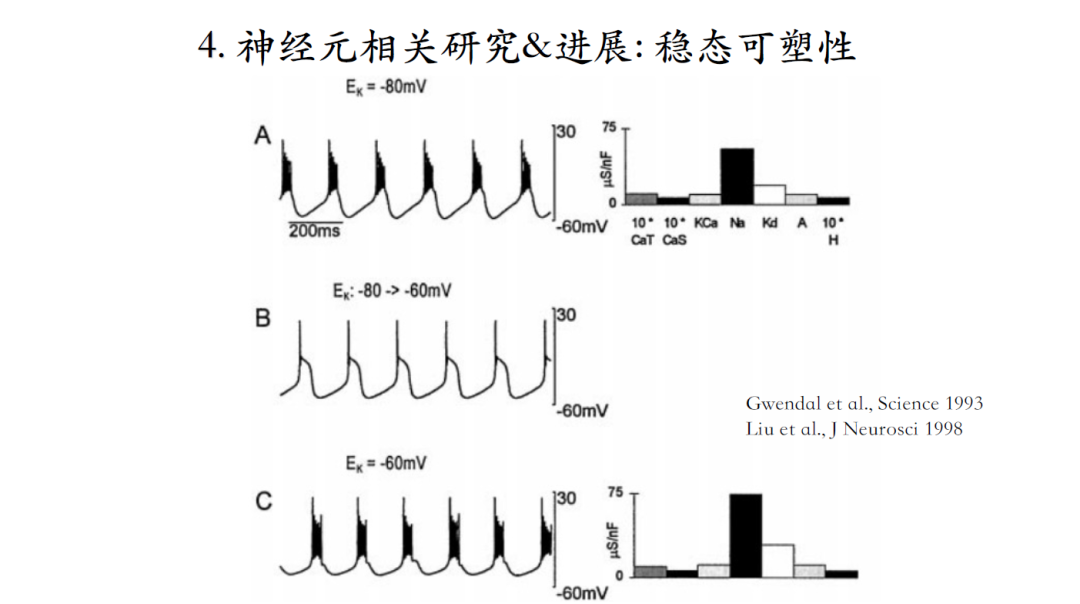
这个模型,可以抵御各种干扰。比如正常生理条件下,钾电流的逆转电势是-80毫伏,如果通过改变细胞外钾离子浓度,使钾电流逆转电势到上升到-60毫伏,这个模型会有一个瞬间的改变,plateau上的脉冲全部消失。经过一段时间之后,虽然恢复了原有脉冲波形模式,但是离子电流的幅值相对于原有模式已经完全不同了。
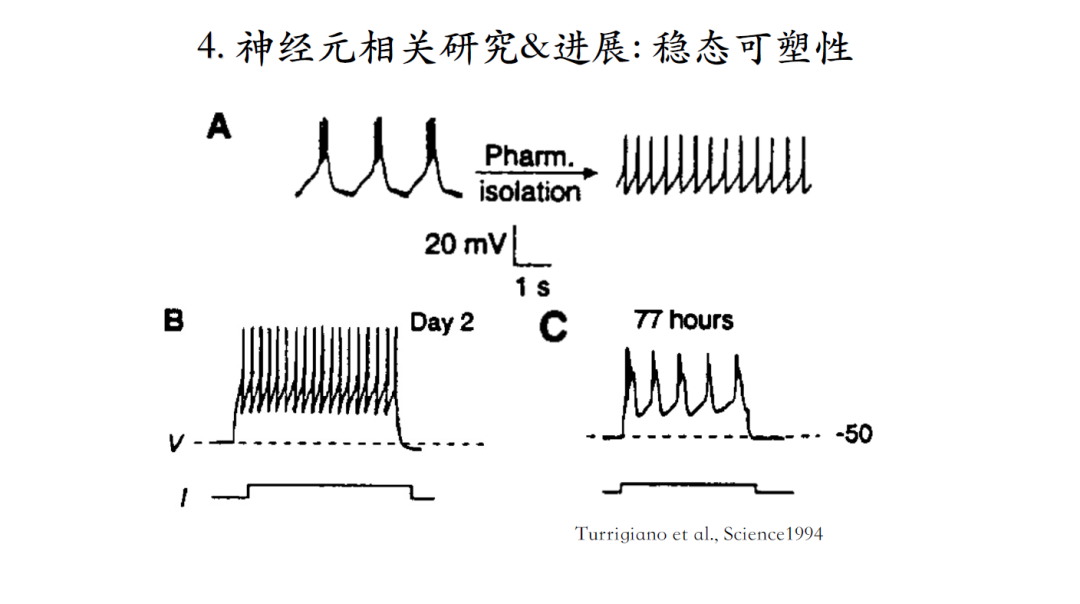
这个理论先有模型预测,然后有了实验验证,作者们通过药物将神经元的脉冲输出模式从bursting改变成为tonic spiking的激发模式。可是77小时后我们可以发现神经元又恢复了原有的脉冲输出模式,进而证实了稳态可塑性模型的机理[20]。
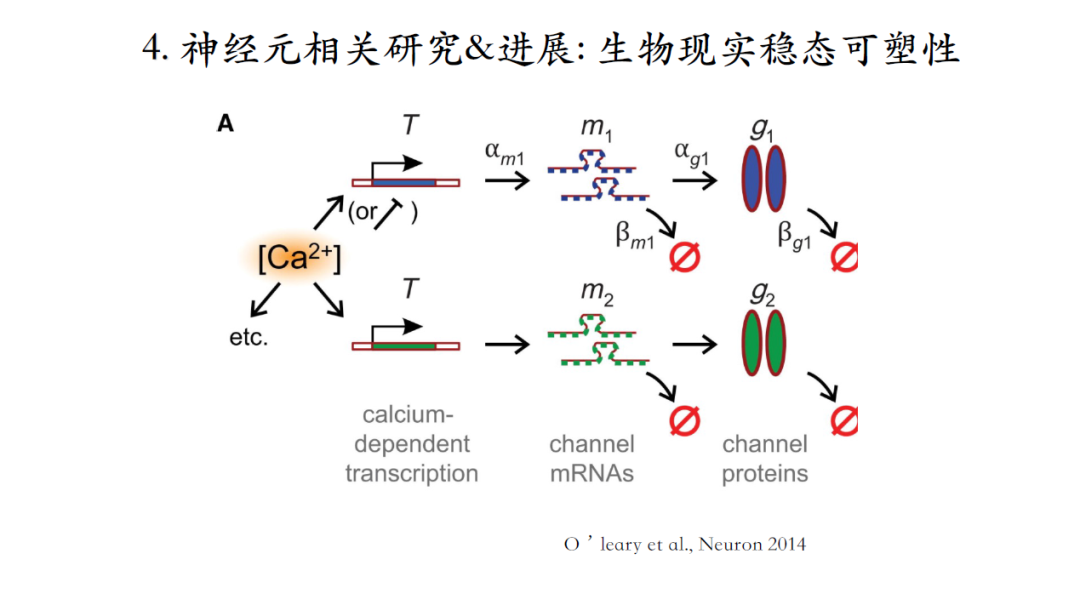
过去的模型缺乏生物实现过程的描述,这个图中模型构建了从DNA到RNA再到最后蛋白质整个生物合成过程的负反馈回路,具有一定生物现实性。
钙离子浓度信号作为信使代表神经元输出的目标模式,如果实时钙离子浓度信号与此目标信号不一致,积累起来的误差信号就会驱动相应离子电流发生改变,直到系统达到稳定的setpoint[21]。
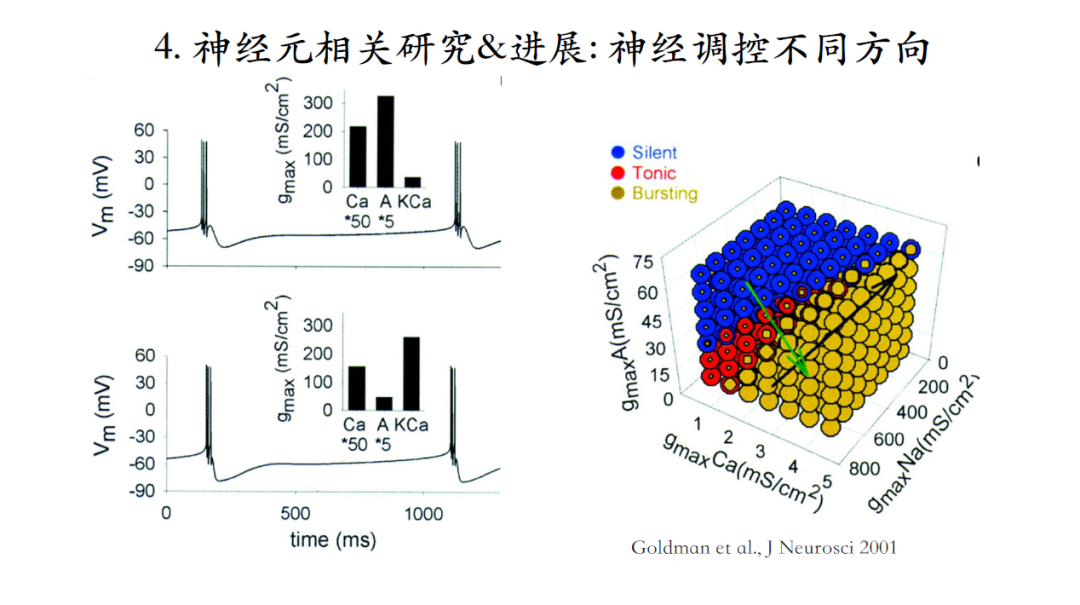
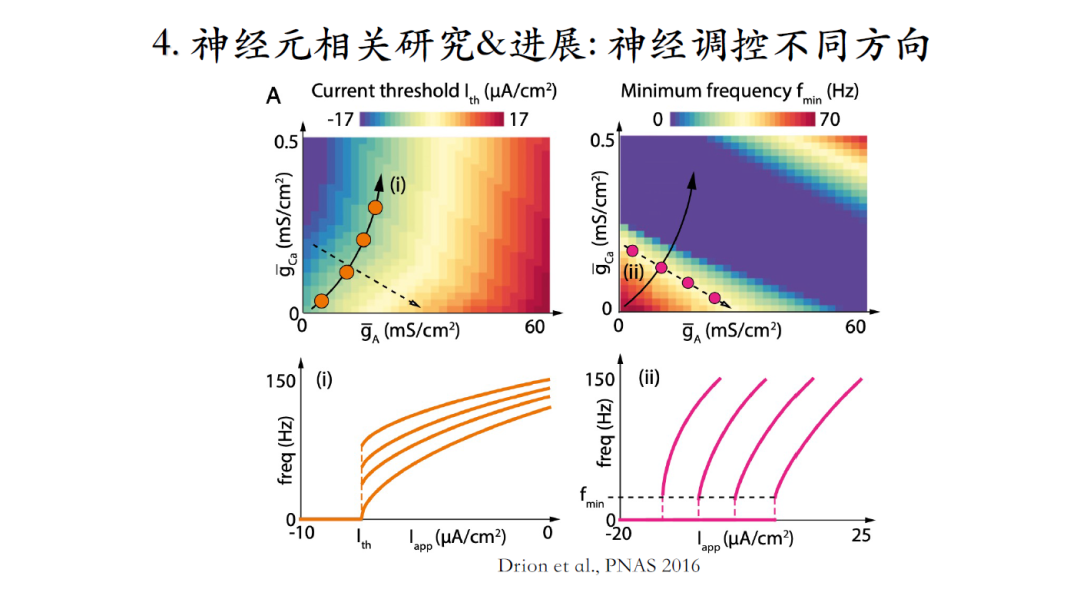
如图所示,神经元接收神经递质调控时,有时会对神经元的输出产生定性改变,有时则不会。神经调质的调控并不具有特异性,会同时影响好几种电流。每个神经元个体又拥有个体差异性,那么接收到神经调质后,什么样的机制可以使具有个体差异性的神经元仍然维持相似的模型输出?
Goldman等人通过搜索神经元模型的三维参数空间,发现黄色区间是bursting空间,蓝色是silent空间,而红色是tonic bursting空间。假如通过神经调质去定性改变神经元的发放模式,应该沿着绿色箭头这个方向让调质去改变相应的离子电流;如果不想定性地改变神经元的发放模式而是想维持当前脉冲输出特性,则需要沿着黑色箭头的方向改变,即不同种类的离子电流通过协同作用可以使神经元在接受刺激后仍然保持稳定的脉冲输出特性[22]。
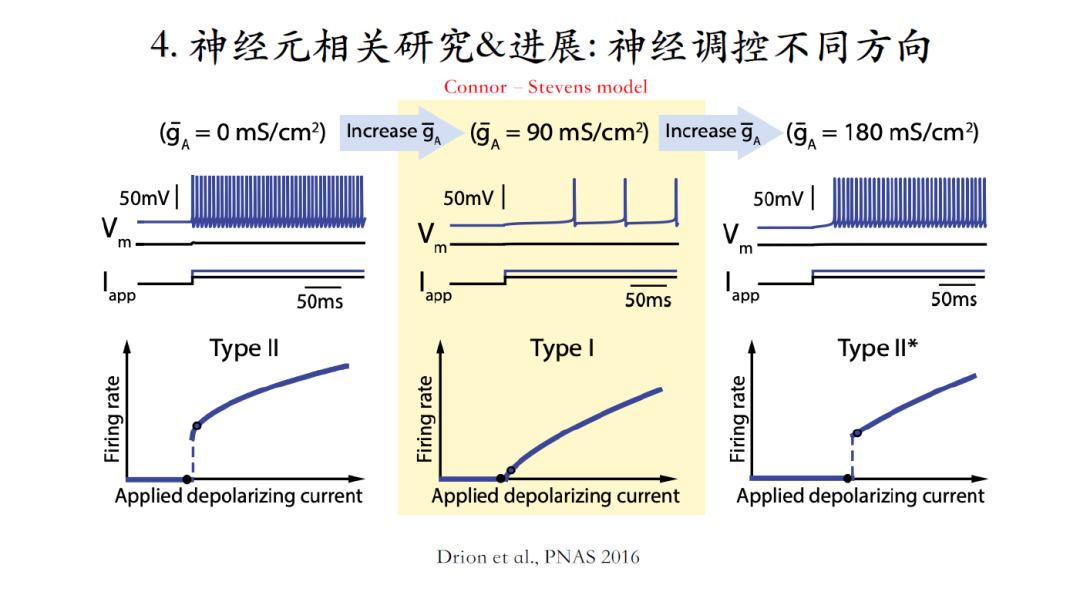
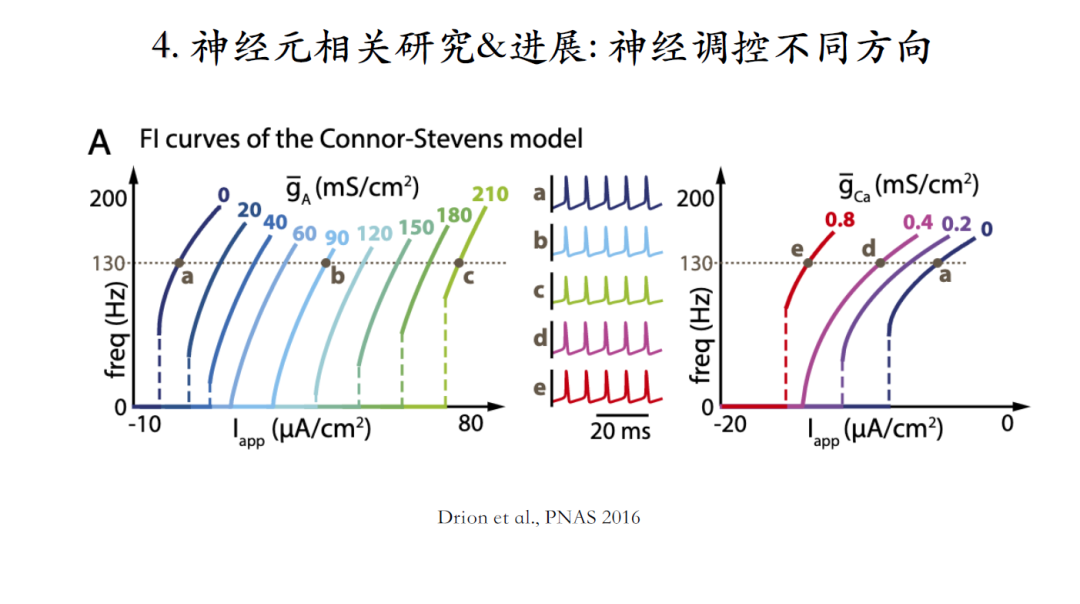
发现一,在type II的Connor-Stevens模型中引入一定量的钾电流可以使其从type II转变成type I,但是引入过量的钾电流又会使其重新变回type I神经元[23]。
发现二,引入钙电流也可以实现同样的转变过程。这些都是一维参数的改变。
如果同时改变模型中两个参数,如上图同时增加A型钾电流和钙电流。发现沿着橙色小点的曲线方向改变可以维持阈值电流保持不变,却会将神经元的最小脉冲频率降低,也即性质上从type II转变成为type I神经元。如果沿着紫色小点的曲线方向改变,可以发现最小脉冲频率没有改变,而极大增加了离子电流的脉冲阈值。因此,在改变多个参数时,可以实现维持一个特性不变的同时,改变另一个特性的目的。
但是当增加一个维度,将参数空间升至3维,会发现type I和type II很难再找到清晰边界,不好实现维持一种特性不变而改变另一种特性的参数变化路径。维度继续升高,个体差异性就体现得更加明显,更难实现参数空间的鲁棒性。
总结说,在低维情况下的一般原则可能是适用的,但在高维空间却不一定适用。如果想要实现上述变化,对于操作单个神经元变化而言可能要求会很高,如何通过约束其他因素增加每一个维度的空间,如在高维空间中增加解的范围,来增加模型的鲁棒性是老师正准备发表的一篇文章。
到目前为止,介绍的工作都是围绕着离子电流密度或者离子通道数量实现,但是稳态可塑性还包括其他方面。
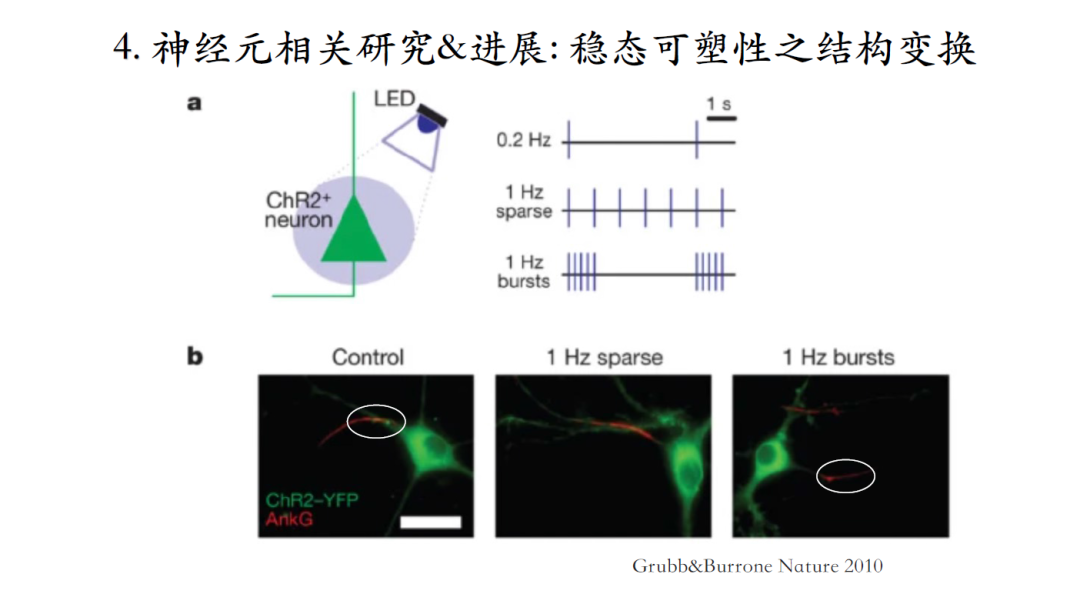
在海马体神经元中引入一个光敏感离子通道,实现人为操纵神经元脉冲发放的目的,会发现神经元轴突初段在经历长时间的burst脉冲激发后会逐渐向远端移动。移动的目的是为了降低神经元的兴奋性(理论上存在争议!),只有降低兴奋敏感度才能使神经元维持一个与初始态相似的发放模式[23]。
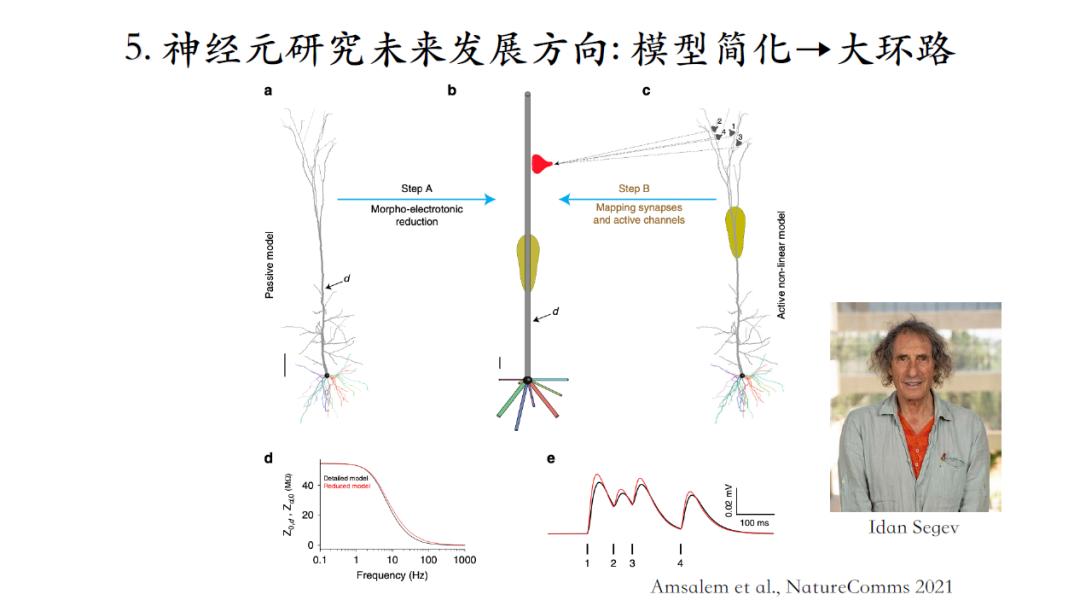
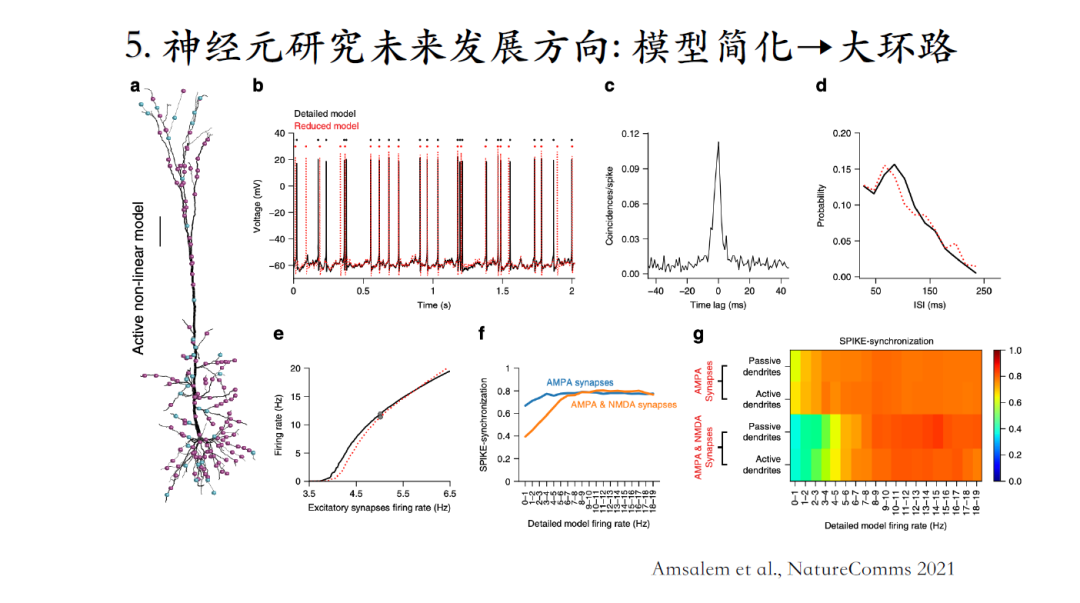
通过Compartment模型做大规模的环路仿真是比较困难的,计算的变量非常多非常复杂,对算力要求极高!诸如Allen Brain Institute,Stanford University,Blue Brain Project等背靠大平台的一些研究者有在开展相关研究。但是也不能无限制的增加模型参数,一个可行的方法是将多个Compartment简化成少数Compartment,简化结构的同时也要保留我们认为比较重要的模型计算特性[24]。
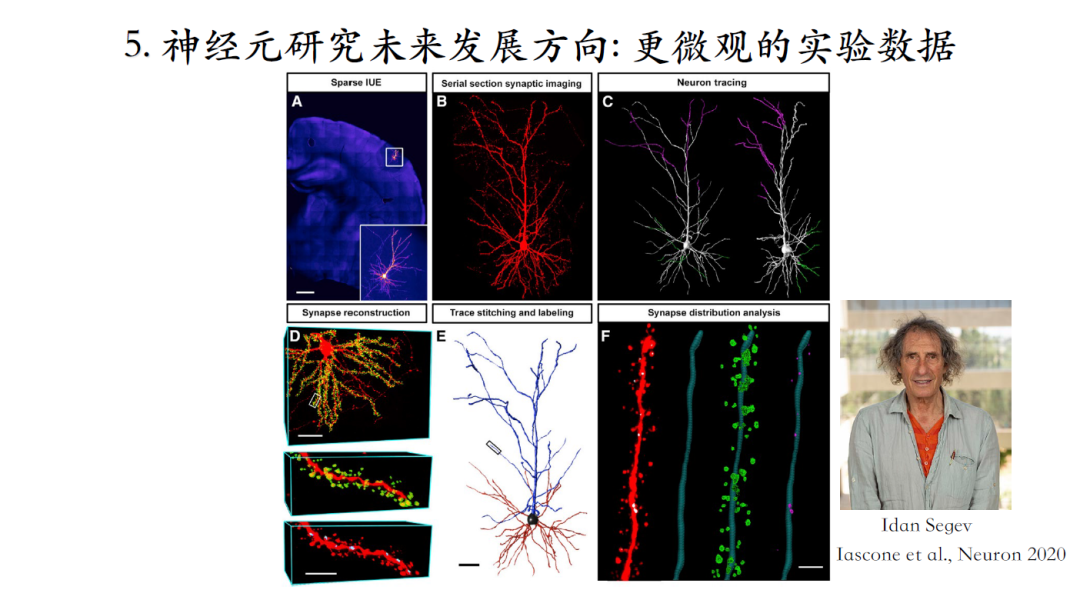
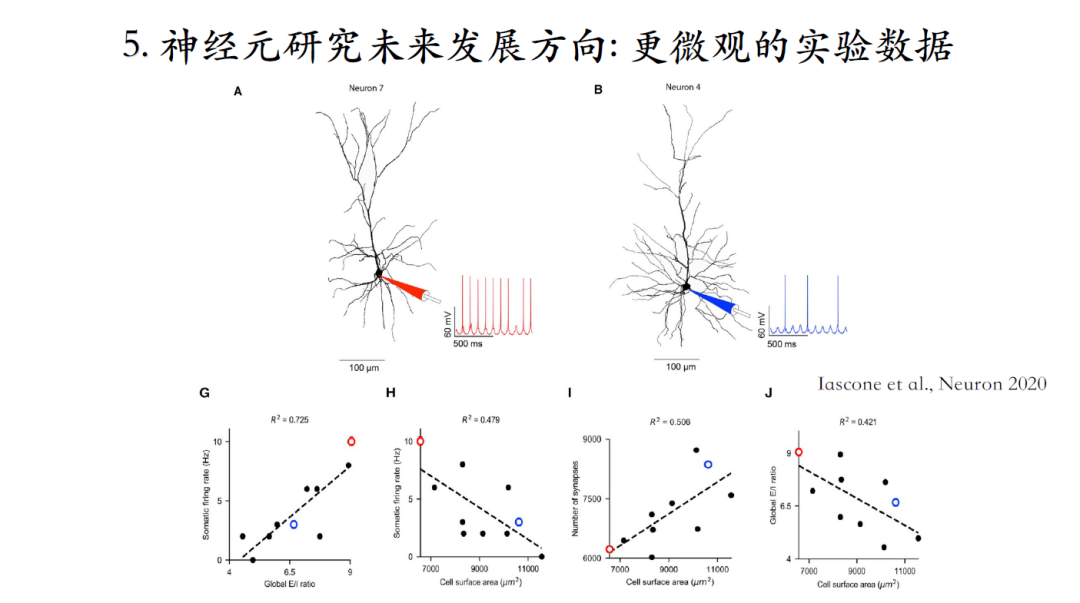
还可以获知更微观的结构信息,例如突触如何分布,兴奋性突触和抑制性突触的比例、分布模式等,结合上述参数可以做更加接近生物现实的仿真。上图中的两个神经元的结构非常相似,可是兴奋性突触和抑制性突触的比例以及分布不同,因此这两个神经元的计算特性也非常不同[25]。
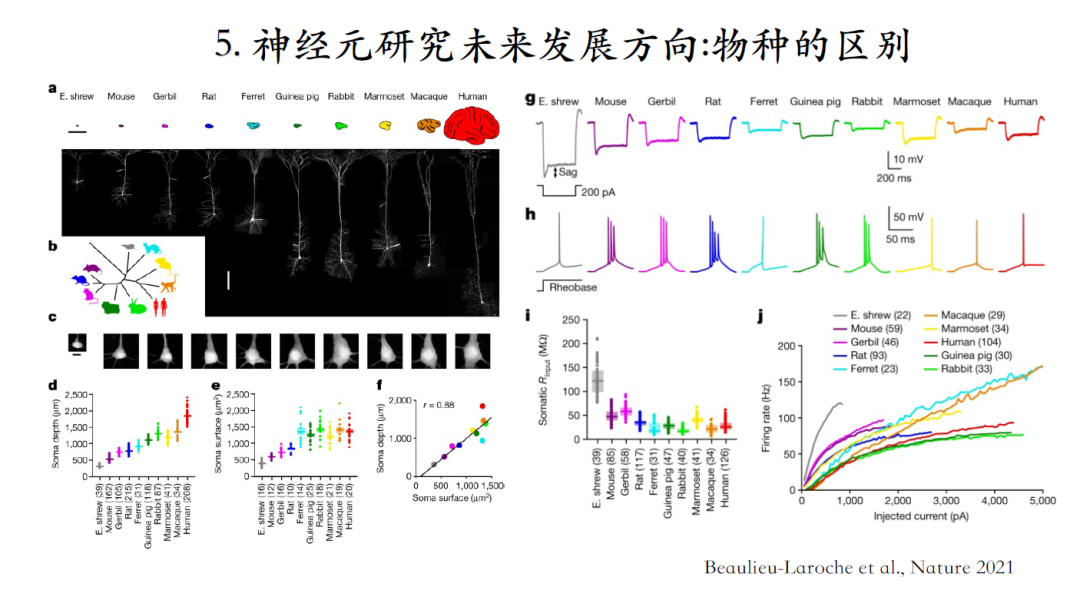
我们一直假设,不同物种和神经元中的一些功能或者计算原理具有普适性。因此我们有时需要从小鼠实验发现中推断人脑的工作机制。
但是最新研究表明,不同物种的神经元从形态到脉冲激发模式都千差万别,很难认为它们实现相同的计算功能[26]。
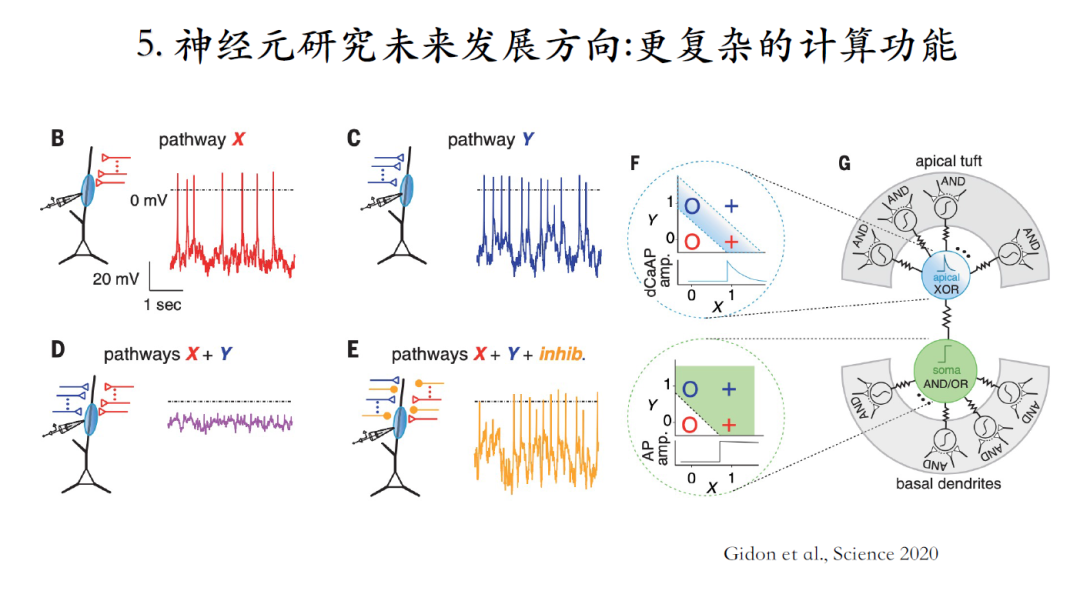
感知机模型中单个神经元处理信息的单层神经网络无法实现XOR门计算。在上述神经元中施加X信号或者Y信号都可以得到对应的脉冲输出,但是同时施加两种信号却没有脉冲输出,即类似XOR门计算[27]。
类脑智能取之于脑却又不应完全受限于脑,因为生物脑的尺寸是有限制的,而机器脑却不受这些限制。想要探索这个领域的人需要极度了解两大领域,一是神经元,二是人工智能。懂第二方面的人比第一方面多,反过来做树突计算的人也不一定了解人工智能领域知识。臧老师建议大家不一定拘泥于神经元的计算原理,可以探索无穷的可能性去进一步提升人工智能的性能[28]。
 [1]Yazan N. Billeh. Systematic Integration of Structural and Functional Data into Multi-scale Models of Mouse Primary Visual Cortex. neuron, 2020, 106(3): 388-403.e18
[2]A. L. Hodgkin, A. F. Huxley. A quantitative description of membrane current and its application to conduction and excitation in nerves. J. Physiol., 1952, 117(4): 500-544
[3]JR Clay. Determining K+ channel activation curves from K+ channel currents often requires the Goldman–Hodgkin–Katz equation. Front. Cell. Neurosci, 2009
[4]W Mel B.. Information processing in dendritic trees, 1994, 6(6): 1031–1085
[5]Laurent Badel, Sandrine Lefort, Romain Brette, et al. Dynamic I-V Curves Are Reliable Predictors of Naturalistic Pyramidal-Neuron Voltage Traces. Journals, 2008
[6]Guillaume Drion, Timothy O’Leary, Eve Marder. Ion channel degeneracy enables robust and tunable neuronal firing rates. proceedings of the national academy of sciences, 2015, 112(38): E5361-E5370
[7]J. C. Eccles, R. Llinás, K. Sasaki. The excitatory synaptic action of climbing fibres on the Purkinje cells of the cerebellum. The Physiological Society, 1966
[8]Jorge Golowasch, Mark S. Goldman, L. F. Abbott, et al. Failure of Averaging in the Construction of a Conductance-Based Neuron Model. JOURNALS, 2002
[9]Zang Y., Dieudonné S., De Schutter E.. Voltage-and branch-specific climbing fiber responses in Purkinje cells[J]. Cell reports, 2018, 24(6): 1536–1549
[10]Michael Häusser. Dendrites: bug or feature?. current opinion in neurobiology, 2003, 13(3): 372-383
[11]Zang Y., De Schutter E.. The cellular electrophysiological properties underlying multiplexed coding in Purkinje cells[J]. Journal of Neuroscience, 2021, 41(9): 1850–1863
[12]Zang Y., De Schutter E.. The cellular electrophysiological properties underlying multiplexed coding in Purkinje cells[J]. Journal of Neuroscience, 2021, 41(9): 1850–1863
[13]Yunliang Zang, Sungho Hong, Erik De Schutter. Firing rate-dependent phase responses of Purkinje cells support transient oscillations. Research Article, 2020
[14]de la Rocha, Jaime, Doiron, et al. Correlation between neural spike trains increases with firing rate. nature, 2007, 448(7155): 802-806
[15]Kuramoto Yoshiki. Cooperative Dynamics of Oscillator CommunityA Study Based on Lattice of Rings. progress of theoretical physics supplement, 1984
[16]Schulz, David J, Goaillard, et al. Variable channel expression in identified single and electrically coupled neurons in different animals. nature neuroscience, 2006, 9(3): 356-362
[17]David J. Schulz, Jean-Marc Goaillard, Eve E. Marder. Quantitative expression profiling of identified neurons reveals cell-specific constraints on highly variable levels of gene expression. PNAS, 2007
[18]Gwendal LeMasson, Eve Marder, L. F. Abbott. Activity-Dependent Regulation of Conductances in Model Neurons. science, 1993, 259(5103)
[19]Zheng Liu, Jorge Golowasch, Eve Marder and L. F. Abbott. A Model Neuron with Activity-Dependent Conductances Regulated by Multiple Calcium Sensors. Journal of Neuroscience, 1998
[20]Gina Turrigiano, L. F. Abbott, Eve Marder. Activity-Dependent Changes in the Intrinsic Properties of Cultured Neurons. science, 1994, 264(5161)
[21]Timothy O’Leary. Cell Types, Network Homeostasis, and Pathological Compensation from a Biologically Plausible Ion Channel Expression Model. neuron, 2014, 82(4): 809-821
[22]Mark S. Goldman, Jorge Golowasch, Eve Marder , et al. Global Structure, Robustness, and Modulation of Neuronal Models. Journal of Neuroscience, 2001
[23]Grubb, Matthew S., Burrone, et al. Activity-dependent relocation of the axon initial segment fine-tunes neuronal excitability. nature, 2010, 465(7301): 1070-1074
[24]Amsalem, Oren, Eyal, et al. An efficient analytical reduction of detailed nonlinear neuron models. nature communications, 2020, 11(1): 1-13
[25]Daniel Maxim Iascone. Whole-Neuron Synaptic Mapping Reveals Spatially Precise Excitatory/Inhibitory Balance Limiting Dendritic and Somatic Spiking. neuron, 2020, 106(4): 566-578.e8
[26]Beaulieu-Laroche, Lou, Brown, et al. Allometric rules for mammalian cortical layer 5 neuron biophysics. nature, 2021, 600(7888): 274-278
[27]Albert Gidon, Timothy Adam Zolnik, Pawel Fidzinski, et al. Dendritic action potentials and computation in human layer 2/3 cortical neurons. science, 2020, 367(6473)
[28]David Beniaguev. Single cortical neurons as deep artificial neural networks. neuron, 2021, 109(17): 2727-2739.e3
[1]Yazan N. Billeh. Systematic Integration of Structural and Functional Data into Multi-scale Models of Mouse Primary Visual Cortex. neuron, 2020, 106(3): 388-403.e18
[2]A. L. Hodgkin, A. F. Huxley. A quantitative description of membrane current and its application to conduction and excitation in nerves. J. Physiol., 1952, 117(4): 500-544
[3]JR Clay. Determining K+ channel activation curves from K+ channel currents often requires the Goldman–Hodgkin–Katz equation. Front. Cell. Neurosci, 2009
[4]W Mel B.. Information processing in dendritic trees, 1994, 6(6): 1031–1085
[5]Laurent Badel, Sandrine Lefort, Romain Brette, et al. Dynamic I-V Curves Are Reliable Predictors of Naturalistic Pyramidal-Neuron Voltage Traces. Journals, 2008
[6]Guillaume Drion, Timothy O’Leary, Eve Marder. Ion channel degeneracy enables robust and tunable neuronal firing rates. proceedings of the national academy of sciences, 2015, 112(38): E5361-E5370
[7]J. C. Eccles, R. Llinás, K. Sasaki. The excitatory synaptic action of climbing fibres on the Purkinje cells of the cerebellum. The Physiological Society, 1966
[8]Jorge Golowasch, Mark S. Goldman, L. F. Abbott, et al. Failure of Averaging in the Construction of a Conductance-Based Neuron Model. JOURNALS, 2002
[9]Zang Y., Dieudonné S., De Schutter E.. Voltage-and branch-specific climbing fiber responses in Purkinje cells[J]. Cell reports, 2018, 24(6): 1536–1549
[10]Michael Häusser. Dendrites: bug or feature?. current opinion in neurobiology, 2003, 13(3): 372-383
[11]Zang Y., De Schutter E.. The cellular electrophysiological properties underlying multiplexed coding in Purkinje cells[J]. Journal of Neuroscience, 2021, 41(9): 1850–1863
[12]Zang Y., De Schutter E.. The cellular electrophysiological properties underlying multiplexed coding in Purkinje cells[J]. Journal of Neuroscience, 2021, 41(9): 1850–1863
[13]Yunliang Zang, Sungho Hong, Erik De Schutter. Firing rate-dependent phase responses of Purkinje cells support transient oscillations. Research Article, 2020
[14]de la Rocha, Jaime, Doiron, et al. Correlation between neural spike trains increases with firing rate. nature, 2007, 448(7155): 802-806
[15]Kuramoto Yoshiki. Cooperative Dynamics of Oscillator CommunityA Study Based on Lattice of Rings. progress of theoretical physics supplement, 1984
[16]Schulz, David J, Goaillard, et al. Variable channel expression in identified single and electrically coupled neurons in different animals. nature neuroscience, 2006, 9(3): 356-362
[17]David J. Schulz, Jean-Marc Goaillard, Eve E. Marder. Quantitative expression profiling of identified neurons reveals cell-specific constraints on highly variable levels of gene expression. PNAS, 2007
[18]Gwendal LeMasson, Eve Marder, L. F. Abbott. Activity-Dependent Regulation of Conductances in Model Neurons. science, 1993, 259(5103)
[19]Zheng Liu, Jorge Golowasch, Eve Marder and L. F. Abbott. A Model Neuron with Activity-Dependent Conductances Regulated by Multiple Calcium Sensors. Journal of Neuroscience, 1998
[20]Gina Turrigiano, L. F. Abbott, Eve Marder. Activity-Dependent Changes in the Intrinsic Properties of Cultured Neurons. science, 1994, 264(5161)
[21]Timothy O’Leary. Cell Types, Network Homeostasis, and Pathological Compensation from a Biologically Plausible Ion Channel Expression Model. neuron, 2014, 82(4): 809-821
[22]Mark S. Goldman, Jorge Golowasch, Eve Marder , et al. Global Structure, Robustness, and Modulation of Neuronal Models. Journal of Neuroscience, 2001
[23]Grubb, Matthew S., Burrone, et al. Activity-dependent relocation of the axon initial segment fine-tunes neuronal excitability. nature, 2010, 465(7301): 1070-1074
[24]Amsalem, Oren, Eyal, et al. An efficient analytical reduction of detailed nonlinear neuron models. nature communications, 2020, 11(1): 1-13
[25]Daniel Maxim Iascone. Whole-Neuron Synaptic Mapping Reveals Spatially Precise Excitatory/Inhibitory Balance Limiting Dendritic and Somatic Spiking. neuron, 2020, 106(4): 566-578.e8
[26]Beaulieu-Laroche, Lou, Brown, et al. Allometric rules for mammalian cortical layer 5 neuron biophysics. nature, 2021, 600(7888): 274-278
[27]Albert Gidon, Timothy Adam Zolnik, Pawel Fidzinski, et al. Dendritic action potentials and computation in human layer 2/3 cortical neurons. science, 2020, 367(6473)
[28]David Beniaguev. Single cortical neurons as deep artificial neural networks. neuron, 2021, 109(17): 2727-2739.e3
为了促进神经科学、系统科学以及计算机科学等多领域学术工作者的交流合作,吸引更多朋友共同探索脑科学与类脑研究,周昌松、臧蕴亮、杨冬平、郭大庆、陈育涵、曹淼、刘泉影、王大辉、刘健、王鑫迪等来自国内外多所知名高校的专家学者在集智俱乐部共同发起「神经动力学模型」读书会,历时四个月研讨,近日圆满结束。
本季读书会形成了聚集500+成员的神经动力学社区,积累了40+小时综述、解读、研讨的视频记录,以及多篇社区成员总结的词条、笔记、翻译、科普资料等。现在报名加入读书会,即可加入社区交流讨论(微信),并解锁相关视频、文本资料。我们对脑的探索才刚刚起航,欢迎你一道参与,共同点亮更多脑科学研究的岛屿!
详情请见:
500+神经动力学社区成员,邀你共同点亮更多脑科学研究的岛屿