Storm, Johan F., et al. “An integrative, multiscale view on neural theories of consciousness.” Neuron 112.10 (2024): 1531-1552.
https://doi.org/10.1016/j.neuron.2024.02.004
心脑问题,即我们的意识体验如何与物质大脑过程相关联,已被哲学家们讨论了几个世纪,并且仍然是科学中最深刻的未解之谜之一。[1-3]过去几十年见证了理论和实证意识研究的激增。各种神经科学的意识理论被提出并受到热议(如Seth & Bayne[4])。通过诸如由Templeton世界慈善基金会赞助的Cogitate联盟[5]等“对抗性合作”模式,不同理论的竞争与测试已然展开。
从历史来看,大部分研究者更倾向于孤立地发展及验证各自所倾向的理论框架。然而,本文则选择了一种别样的路径。它源起于在与意识相关的主题上工作的欧盟“人脑计划”(HBP;2013-2023)内的研究人员,他们代表着不同的理论观点。在近期的实证研究进展基础上,我们发现声称相互矛盾的各理论,在解释意识的不同方面时,往往在基本的神经机制与过程上展现出趋同之态。在这里,我们认为几种理论至少在某种程度上是兼容和互补的,并且我们考虑了迄今为止被很大程度上忽视的趋同方法。
鉴于无法涵盖所有提出的基于神经科学的意识理论,我们在此重点介绍五种我们认为具有代表性且互补的理论:(1) 全局神经工作空间理论(GNWT[6]),(2) 整合信息理论(IIT[7]),(3) 循环处理理论(RPT[8]),(4) 预测处理( PP[9-11])和神经表征主义(NREP[12-13]),(5) 树突整合理论(DIT[14])。这些理论涵盖了Seth和Bayne[4]概述的四大类意识理论中的三类,因此可以广泛讨论如何调和和整合不同的理论。
在这里,我们使用广义的“理论”(theories)一词,因为该术语没有公认的定义(见表1),其他术语(“假设”,“模型”和“框架”[hypothesis, model, framework])也缺乏严格的定义。我们关注这些理论和观点的内容,而不是它们的地位或成熟度。我们选择理论时使用的另一个标准是,每个理论都应代表一个在可定义的思想框架下汇集的神经科学研究的累积体。
通过比较这五种理论以及如何部分融合,我们旨在启动一种整合过程,未来可能还会融入其他理论中的有价值元素。然而,当前的一些理论可能被证明是完全或大部分错误的或无效的,并将被未来的选择所淘汰,正如科学史上反复发生的那样。为了解决长期存在的、看似无法解决的哲学和理论争论,我们提出了一种不同的方法,关注不同层面上基于经验的整合、复杂性、表征和不同层次的循环处理的概念,这些概念似乎是这些理论的共同特征。
根据Ned Block的观点,我们将现象意识(PC,即时的主观体验,如看到蓝天的体验)的机制与访问意识(AC,理性控制的意识)和可报告性机制区分开来。我们讨论了这些区分如何与提出的意识实证测量方法(背景知识1和2)相关,以及它们如何与注意、期望、计划行为和不同形式的记忆相关[8],[12],[25],[26],[27],[28]。我们还区分了从低层次的机制特征到高层次的系统属性的解释层次。
背景知识 1:评估意识状态和水平的方法
识别能够可靠地指示人类意识存在与否的基于大脑的测量方法,一直是大量研究文献的焦点,且为临床医生所关心,可谓是理解意识及其物理基础关系的重要起点。在行为评估意识的可靠性受到质疑的情况下,如全身麻醉期间或因严重脑损伤而处于无反应状态的受试者,基于大脑的测量方法显得尤为必要。
第一类标志涉及绕过运动行为,直接测量大脑对感官刺激(如口头命令或复杂的声音序列)的反应。此类标志的一个突出例子是P3b,它由听觉序列中整体违反预期引发。根据全局神经工作空间理论,P3b表明感觉信息已广播至额叶皮层处理器,已用来检测脑损伤患者的意识存在。虽然P3b提供了一个特定的标志(即,它仅在意识到刺激的受试者中出现),但它在有意识的脑损伤患者和健康的有意识受试者中经常缺失[30],[31]。P3b的这种低敏感性提醒我们,访问意识的神经相关物(指示与环境的连接性和/或高层次认知功能的参与)与现象意识的相关物之间可能存在分离。
第二类标志直接关注脑活动,不必依赖于感官处理和认知功能。最近,关于整合与分化的测量方面出现了实证趋同,展示了更准确地检测意识的可能。值得注意的是,这种趋同涉及众多独立研究者,他们分别支持不同的理论框架,并在多样的条件下使用不同的技术,如睡眠、梦境、麻醉、癫痫、幻觉状态及昏迷等[32]。一般来说,这种可靠地检测意识的方法,独立于外部环境的连接性、行为表现、认知和记忆能力,不仅具有临床意义,也推动了对现象意识神经机制的深入探索。复杂性测量要么依赖于自发大脑动力学的表征(通过功能连接网络和图论指标的分析)[33],[34],要么依赖于直接引发的交互模式的量化皮质扰动。[35]后者以扰动复杂性指数(PCI)为例,在具有挑战性的临床条件下表现出高度的敏感性,并指向大脑复杂性和反馈互动之间的机制联系。事实上,在各种生理和病理条件以及实验模型中,扰动复杂性指数在意识丧失和恢复期间的减少和增加都与循环激活波的消失和重现密切相关[36],[37],[38],[39]。
背景知识 2:评估意识内容的方法
在细粒度的子类别水平上实证评估意识内容,因需准确确定代表特定特征的记录位置,而显得颇具挑战性。Koch等人提出了一种行之有效的方法,当探索意识的特定内容的神经相关性(NCC)时:(1)首先在映射步骤中,使用具有特征特异性的刺激来确定受试者记录位置(如神经元记录中的神经元或功能性磁共振成像研究中的体素)的反应特性;(2)其次,应用一个实验范式,如多稳态图形或双眼竞争,其中物理刺激的特性保持不变,而意识内容在对刺激的不同解释间切换。如果在第一步映射中特征动态活动的变化反映了体验(报告)的意识内容变化,那么相应的记录位置就有可能是潜在的意识的神经相关性;(3)通过实验尝试排除已知的混杂因素,例如,神经活动与意识感知共变可能反映了意识的前提条件或后果[40]。因此,必须记住,这种方法是相关的,而不是因果的,注意和记忆效应可能混淆结果的解释。
这种方法已经成功应用于猕猴和人类的电生理记录。人类颅内(立体脑电图)记录导致了强直活动模式的识别,即对感觉刺激的长时间低振幅伽马带反应,这可能对应于先前报道的反映感知意识的晚期脑电图成分的颅内相关物[41]。躯体感觉刺激在双侧顶叶覆盖区和后岛叶皮层中引发了持久活动。此外,视觉-触觉刺激特别增强了这些稳态成分,这与视觉-触觉刺激显著改善触觉消失患者的检测能力的观点相一致[42],[43]。
这种方法可以扩展到具有足够高空间分辨率的人类功能性磁共振成像(fMRI)测量,以区分皮层柱水平的特征特异性反应。在 Schneider等人的一项研究中,映射阶段定位了在人类中颞中区(hMT+)中选择性响应水平或垂直运动刺激的柱状簇[48]。在主要实验中,呈现了一个模糊的表观运动刺激(运动四重奏),它可以被感知为水平或垂直运动。通过跟踪预先映射的水平/垂直柱状簇的活动,可以预测受试者在交替时间段感知到的双稳态运动刺激(水平或垂直运动)的两种可能解释中的哪一种。运动四重奏研究的结果支持了hMT+是(部分)运动方向内容特异性的神经相关性观点,这与Salzman和Newsome在非人类灵长类动物中阐明的中颞区(MT)在运动视觉中的因果作用一致[49]。更普遍地说,在具有亚毫米级超高场fMRI的模糊感觉刺激期间,映射出皮层(柱状)功能簇并测量其反应符合Chalmers[15]和Koch等人[16]概述的内容特异性神经相关性搜索标准。
在对比众理论时,我们首先需要澄清关键概念、定义所用术语,并确定这些理论试图解释的现象(explananda)。我们的比较方法立即突显的一点是,不同的理论往往关注“意识”的不同方面或子类型,这可能导致概念混淆和误解。相反的理论可能会提倡不同的实验方法,这些方法并不直接可比,对数据的解释亦各不相同。
例如,全局神经工作空间理论主要探讨访问意识(AC)[19](表 1),主要试图解释主体如何访问和利用意识内容,以及意识与无意识的“信息处理”(与现象体验相对),通常研究对特定感官输入的反应,如看到一张脸。相比之下,整合信息理论、循环处理理论和预测处理/神经表征主义,则主要关注现象意识(PC)[19],即体验某事“感觉如何”[18],而不考虑其认知功能如何以及是否可以报告。[50],[51]
因此,与全局神经工作空间理论相比,其他理论(见下文)主要试图解决“难问题”,[1]即解释现象意识和“感受质”如何在类似大脑这样的物理系统中产生,以及为什么这些主观现象会在无梦睡眠或麻醉期间消失(但参见 Sevenius Nilsenet al.的研究[52])。
这种观点上的差异,部分源自于各自独特的哲学立场:与整合信息理论、预测处理/神经表征主义或循环处理理论相反,全局神经工作空间理论的支持者通常不认为现象意识或感受质是一个与访问意识不同的真实现象,而是认为,只要理论能够解释可观察的事实——比如有人报告说:“我现在看到了脸!”——便无需进一步解释。
对于预测处理/神经表征主义,整合信息理论、和循环处理理论,现象意识是主要的解释对象,它们将访问意识内容视作次要,仅作为意识内容应用于运动或认知操作执行过程的辅助。[7],[12],[13],[27],[53]
树突整合理论提出了针对访问意识和现象意识的机制。[54]它主要在细胞和回路层面上运作,并试图解释为什么哺乳动物的意识依赖于皮质-皮质和丘脑-皮质回路,认为访问意识和现象意识的体验涉及这些回路的同时活动,由树突处理所引起。[57],[14],[54]
全局神经工作空间理论与整合信息理论的原始版本还有另一种不同之处:前者主要关注选择和使用访问意识的过程,而后者则更多地关注现象意识的基本性质和特质(就信息整合和交互而言),以及现象意识的状态和水平。[55]然而,现在这两种理论都越来越多地试图解释过程、内容和状态/水平,[28],[16]这一趋势与预测处理/神经表征主义、循环处理理论和树突整合理论的方向一致。[12],[14],[56]越来越多的人认为,任何完整的理论都应该解释意识的内容和状态/水平。
在本综述中,我们将首先简要介绍这五种理论的核心观点,随后探讨它们如何相互比较,借助于大脑的层次结构作为分析的支架,并进一步考察不同理论间是否以及如何可以相互调和,甚至可能实现整合。
全局工作空间理论(GWT)[58]将产生意识体验的处理步骤描述为本地处理器和全局工作空间之间的动态交互,全局工作空间整合信息,并将其广播到其他处理器以进行进一步处理。因此,全局工作空间充当了信息传递的主干,通过它,多个大脑区域得以共享不同感知与认知过程中的信息。被广播的信息,便构成了意识的内容。
后来,受神经生物学启发的全局神经工作空间理论(GNWT)被提出。[6],[28],[59],[60]它采用了全局工作空间的相同功能概念,但将其与特定的脑网络和神经元特性联系起来。此处的局部处理器指那些具备感知、动作、记忆等专长的模块化皮层区域,其活动由感觉刺激所激发,能够独立完成计算任务。举例来说,具有这些特性的模块广泛存在于早期感觉皮层中,不同的特征在专门的皮层区域中得到处理。[61],[62],[63]
全局工作空间涉及的是连接广泛的大脑区域网络的长程连接(皮层-皮层和丘脑-皮层),特别是在前额叶皮层、后顶叶皮层和相关区域。这些分布区域可以共同选择一个心理对象,从而增强和绑定该对象的所有相关特征的表征,并在感知中形成一致的对象表征。这种一致的对象表征对应于知觉心理学中所讨论的基于对象的注意。在这个方案中,丘脑和其他皮层下结构如基底神经节也可以放大和分发信息到各个皮层区域,并整合大脑各大区域的神经元活动。皮层区域被认为可以选择和整合信息,使大脑能够集中注意力并进行复杂的认知任务。最后,当局部神经元集合达到进入全局工作空间并参与后续广播的活动阈值时,这个时刻称为“激活”。
激活是全局神经工作空间理论的一个核心概念,它与前顶叶皮层中的循环处理有关。[64]近年来的研究支持了这一观点,[28]例如,人类神经生理学和神经影像学研究显示,在刺激开始后大约200到300毫秒内,大脑活动的明显分歧依赖于有意识的感知,[65-66]这种现象在不同的感觉模式和实验范式中都有观察到。在最初200毫秒的早期感知处理中,有意识(报告)和无意识(未报告)的试验可能相似甚至相同,[67-69]但有意识的感知与后期事件相关,如人类头皮事件相关电位的N2/P3b/P300成分。[6],[70-72]在某些条件下,有意识访问的时间可以延迟,例如在注意力分散或双任务情况下,全局工作空间已经被另一个有意识对象占据。[67],[73]人类前颞叶的概念细胞和非人类灵长类动物的前额叶神经元在相应刺激被有意识地感知时表现出更强和更持久的放电率。[74-77]在猴子中,信号传播到前额叶皮层进一步导致激活和行为报告。[64]虽然这些实验结果有些存在争议,[12],[16],[78] 通常是因为感知报告可能混淆了后来的额叶活动。[42]然而,最近在猴子中的无报告范式的结果,可能将减轻这些质疑。[79],[80]
脑区内或跨脑区的循环回路支持持久活动和类似激活的过程。[20]全局工作空间所需的长程皮层投射起源于皮层II/III和V/VI层,在这些层中反馈效应也最为显著。[81-84]最近的证据表明,持久神经元活动的回响环路涉及皮层-皮层相互作用和丘脑和小脑核等皮层下区域,[85-86]这与支持全局工作空间的广泛的长程连接网络一致。[87]
(A)在GNWT中,单个处理节点(以圆圈表示)在未与广泛的远程连接网络——即“全局工作空间”(以红色表示)建立连接前,仍处于访问意识之外。
(B)当本地节点通过“激活”过程在被称为“全局工作空间”的广泛远程连接网络(红色)中建立连接时,它们确实会产生(访问)意识。从未连接到全局工作空间的节点仍然处于潜意识和无意识状态(以蓝色节点表示),而已连接的全局工作空间内的过程可供其他认知过程使用。当前未连接到全局工作空间但可通过注意力增强其信号的(“注意力门控”)的方式连接的组件被称为“前意识”(橙色节点;类似于其他一些理论所称的现象意识;见图3、4、5、6和7)。节点之间的非活动连接用细黑线表示,活动连接用粗(彩色)线表示。
整合信息理论(IIT)通过强调经验的“现象结构”,使用内省来描述现象属性,然后根据相应的物理属性推导这些属性。这种方法允许通过测量行为、功能和神经相关性来验证或证伪这种物理解释。[50]这种以意识为先的思路的种子可以在1998年发表的早期论文“意识与复杂性”中找到。[88]在这篇论文中,作者从内省出发,假设每一个意识体验既是整合的(形成一个统一的整体)又是高度信息化的(每个意识状态与大量的替代状态有明显不同)。他们随后假设,神经元的整合和分化的共存,是意识系统的关键物理属性,并将这一假设操作化为可测量的神经元属性:
1. 整合:一组神经元只有在成为功能簇的一部分,通过数百毫秒内的持续交互实现高度整合时,才能对意识体验做出贡献。
2. 分化:为了维持意识体验,这个功能簇必须能够拥有大量不同神经状态的组合,并与高复杂度值相关。
这种将意识与神经元交互的复杂性联系起来的原则,构成了IIT早期论述的核心,[89],[90]它启发了“弱IIT”的版本,[91]并衍生出一系列可测试的解释和预测:
1. 一些具有适当解剖要求(高密度的侧向连接和大量折返环路)的丘脑-皮质系统区域支持意识体验,而具有模块化或前馈架构的其他结构(如小脑)则不支持。
2. 同步或广泛的神经元激活不一定与意识状态相关,除非它们具有高水平的分化或复杂性。
3. 在解剖要求得到保留的情况下,功能的改变影响折返、整合和分化,可能会在睡眠、全身麻醉和全身性癫痫等情况下导致复杂性和意识的丧失。
4. 即使在没有感官输入、运动输出和执行功能的情况下,高复杂性和意识体验也可以由内在的大脑互动支持。
5. 通过直接的皮层干预和记录,可以测量脑内因果互动的复杂性,从而可靠地评估意识的存在与否。[92]
在过去的20年里,这些早期预测已使用各种脑成像模式在不同的意识状态(清醒、睡眠、做梦、迷幻状态、癫痫和意识障碍)中进行了广泛评估,并显示出临床应用的前景(综述见 Sarasso[32];另见[背景知识1])。整合信息理论的完整表述[93],[94]在其4.0版本中达到高峰,[95]它考虑了意识的五个现象属性(内在性、信息、整合、排他性和组合),然后将它们表述为物理属性。通过其数学形式,整合信息理论旨在不仅解释意识的存在与否,还致力于阐释其质量和内容。具体而言,整合信息理论主张每一种体验(特定的“现象结构”)与物理基质所规定的相应“因果结构”之间具有解释性同一性。[50]
整合信息理论认为,因果结构完全捕获了系统在特定状态下的因果能力——例如,一些活跃和一些不活跃的神经元网络。要测试这一理论框架,需结合系统的干扰、观察与分区操作,以精确评估元素子集如何对彼此产生差异并从彼此中获得差异。
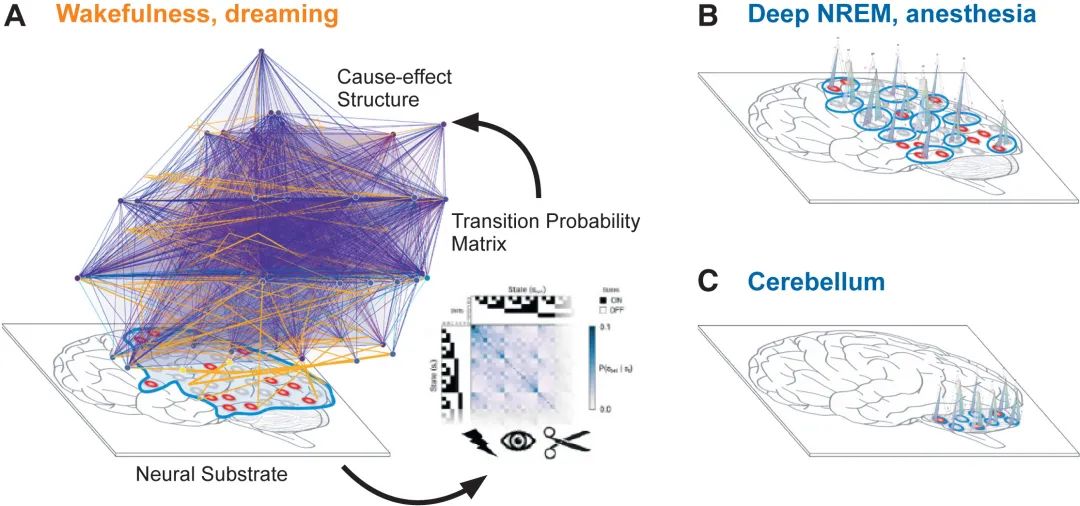
图2.整合信息理论揭示了三种“类脑”基质的因果结构。
(A) 在清醒或做梦期间,后皮层在给定状态下由高度集成的网络组成的神经基质(红色表示活跃神经元,灰色表示沉默神经元)。通过系统扰动(闪电标志)和观测(眼睛标志),可以获取系统的转移概率矩阵(TPM)。这种TPM通过分割操作(剪刀标志)来评估不可约的因果机制及其相互作用,从而揭示内在的因果力量,展现一个具有高度复杂性(高Φ值)的因果结构,充满了多样的区分与联系。
(B) 解剖学连接相同,神经调节和突触后特性的变化可能导致因果相互作用的崩溃。因此,高 Φ (A) 的单个大因果结构“分解”为多个不相交的结构(灰线的“峰”),每个结构的Φ都很小。如果神经基质的特征是模块化和/或主要是前馈连接,类似于小脑中发现的连接,则网络指定单独的、最小的因果结构,每个结构都具有非常低的Φ。
尽管在计算上,即使是简单的物理基础的因果结构也难以完全展开,但整合信息理论做出了有趣的预测,这些预测是可以通过实证研究进行测试。例如,整合信息理论认为 (1) 具有侧向和循环收敛-发散连接的分层组织的网格状皮质结构(如在后皮质中发现的那些)最适合整合信息,因此对应于意识的神经基质;[96]并且 (2) 现象空间——延展性的感觉——可以通过皮质区域的网格状结构所规定的“因果结构”来解释;[97] (3) 在这些网格内,不活跃的神经元也像活跃的神经元一样对因果结构和体验做出贡献;(4) 最后,模块化或前馈架构(如深度学习网络)即使其可观察到的表现等同于有意识的人类,也不会有意识。[94]
循环处理理论(RPT)的核心思想是循环(反馈,回返)处理(RP)对于意识体验是必不可少的,甚至可能是充分的。Victor Lamme基于对灵长类动物视觉的实验[98-100],发展了循环处理理论,并提出了一个关于视觉意识的一般理论[27],[101],[102]。循环处理理论的一个关键元素是它将注意力和意识视为独立(正交)的过程(图3,上方水平箭头和左侧垂直箭头)[8]。这使得循环处理理论与许多其他理论(包括全局神经工作空间理论、预测处理理论和注意力图式理论[103])有所不同,但又与神经表征主义并无区别。
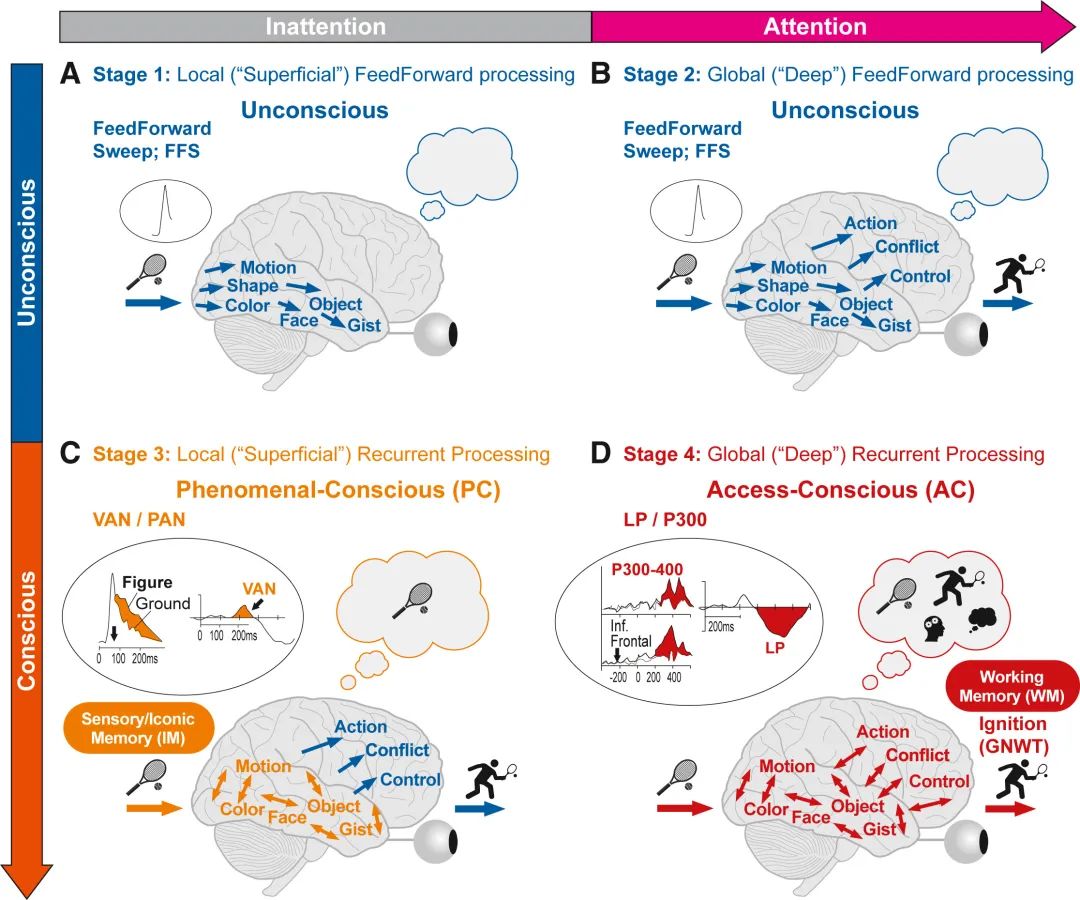
(A-D) 灵长类动物(人类)视觉系统中前馈(FF)和循环(RP)感觉处理的四个阶段(1-4),是两种意识形式的基础;现象意识(PC)和访问意识(AC)。由于 RPT 侧重于视觉,因此该图仅说明视觉处理,尽管RPT假定的一般原则(阶段1-4、PC与AC等)也适用于其他感觉模式(参见Dembski 等)[43]。(A–D)阶段 1–4:请参阅正文中的循环处理理论进行解释。(A和B)显示了早期无意识处理的两个阶段(蓝色)。
(C和D)显示现象意识(C 中的橙色)和访问意识(D 中的红色)处理阶段。椭圆形插图 I(C)和(D)显示了电生理 (EEG/ERP) 成分的示例,这些成分似乎反映了第3阶段的处理(C、VAN:视觉意识消极性;PAN:感知意识消极性[43]),第4阶段处理(D、P300-400和LP(晚期阳性)是晚期事件相关电位(ERP)组件,根据RPT假定它们与全局循环处理相关,并且根据 RPT 假定与全局工作空间的“激活”相关。GNWT(见图 1B),说明了这两种理论之间的经验和机制一致性。第 3 阶段处理的现象意识内容仅短暂(0.1-1 秒)存储在大容量感觉记忆中(对于视觉,称为图像记忆,IM;C)中,而 AC 内容在低容量工作记忆中存储时间更长(>1 秒)(WM;D)。上面的大箭头(A 和 B)说明了注意力与无注意力是一个单独的维度,与有意识/无意识正交(左,垂直箭头),根据 RPT 改编自 Lamme [8],[27],[102] 以及 Koivisto 和 Revonsuo [104],[101],[102],[105]
循环处理理论概述了视觉处理的四个阶段(图 3A-3D):其中前两阶段属于无意识处理,后两阶段则涉及有意识的体验,但二者各异,分别关联现象意识与访问意识。[19]循环处理理论还将现象意识和访问意识与注意力和不同形式的记忆联系起来(图3C和3D),从而开创了将循环处理理论与其他理论统一的尝试[8],[25],[105]。
第 1 阶段——局部(浅表)前馈处理:一种快速的前馈波(FFS,约100–200毫秒)携带信息从初级视觉皮层传递到更高的视觉皮层,在那里提取图像特征。然而,如果没有注意力的增强,处理将停留在局部,无法进入第2阶段,仅产生“阈下”处理[106],不涉及现象意识或访问意识[27],[102],[105](图3A)。然而,FFS可以在所有皮质层引发“非前馈”的循环、侧向或水平处理,从而触发第3和第4阶段。
第 2 阶段——当刺激物受到注意时,发生更全局(“深层”)的前馈处理。在这种情况下,增强的FFS会传播到运动和前额叶区域,导致对主要内容的快速识别(约200毫秒)。这可以触发即时的,无意识的(自动)行为,例如击打网球(图3B),[27],[102]而主体可能对刺激物及其反应并无意识。
第 3 阶段——局部(浅表)循环处理是一个中间处理阶段,其中未被掩蔽的刺激物,主要在腹侧视觉通路中,[107]引起现象意识体验[19]。然而,如果不引起注意,循环处理受限,无法与前额叶和运动区域进行互动。有限的循环意味着感觉刺激不会进入工作记忆和报告所需的进一步认知过程。因此,存在现象意识而没有访问意识(图3C)。
第 4 阶段——在足够的时间下,受到注意的刺激物会导致深层、全局的循环处理(图3D),[27],[102],[105]达到更高的水平,包括前额叶和执行区域,为大脑的执行系统提供可访问的内容(除了现象意识之外的访问意识),用于工作记忆、报告和其他行为反应,例如在全局神经元工作空间理论中的“激活”(见上文)。
在循环处理理论中,注意力对于确定哪些刺激物会导致全局(第4阶段)循环处理和访问意识,而哪些会导致局部(第3阶段)循环处理和现象意识是至关重要的。只有在有注意力的情况下,主体才“知道”并且可以使用和报告其体验(访问意识)。循环处理理论还声称,不同形式的短期记忆与现象意识和访问意识相对应。而全局循环处理会涉及稳健的、低容量的工作记忆(WM),局部循环处理会涉及大容量、非常短暂的感觉(图像)和脆弱记忆(SM,FM;0.1秒到几秒),在感觉皮层中很容易被其他刺激物覆盖[105]。这解释了无意视盲和变化视盲现象,以及为什么在没有提示的简短展示后被试只能报告少数视觉项目(在 WM中保存)[108]。
作为由实验主义者开发的理论,循环处理理论坚定地植根于实证观察,与更多的哲学理论形成对比。许多实验结果支持循环处理理论,例如来自掩蔽、变化视盲、无注意、注意力闪烁、忽视和消失的结果,[8],[101],[102],[105]但并不意味着其他理论就是不对的。早期(<200 毫秒)的事件相关电位(ERP)成分(视觉意识负性;VAN)似乎(在时间和拓扑上)与局部循环处理(第3阶段[104])匹配,并可能反映现象意识(图3C)。晚期(约300毫秒)阳性(LP)事件相关电位(P300/P3b[29])显示出第4阶段的特征,因此可能反映访问意识(图 3D),[102],[105]与全局神经工作空间理论一致[28]。类似的早期事件相关电位也在其他感觉模态中发现,表明一种反映感觉现象意识的普遍感知意识负性(PAN)[43]。
循环处理理论的一个挑战是,循环处理“是否足以形成意识体验[8],[102]”。虽然通过区分现象意识和访问意识可以解决几项看似驳斥循环处理理论的实证挑战,[8],[19]但核心问题仍然存在:循环处理似乎远比意识普遍,这表明单靠循环处理并不足以形成意识[102]。实际上,在明显无意识的状态下(无梦睡眠、麻醉、昏迷)循环处理并未完全消失[102] [52]。Lamme提出“缺失的成分”可能是神经可塑性,因为循环处理可能满足赫布法则,从而“根本区别于”前馈处理[8],[109]。因此,意识处理可能用于促进感知学习[101],[102]。
预测处理(PP)的关键思想,更具体地说是以神经表征主义(NREP)作为意识理论,提出了一个与众不同的视角:大脑并非简单地从外部世界提取或复制知识以实现感知,而是通过构建一个内部的世界模型来建立感知和知识。这一构建过程是通过推断当前接收到的感觉输入的最可能原因来实现的。因此,所谓的“预测”并不一定是指向未来的预测,更多是指对“此时此地”的现象进行推断。通过比较内部表征的效果与实际感觉输入,可以计算出两者之间的预测误差,这既有助于感知学习,也有助于推理。当特定情况在皮质-丘脑系统中实现时,推理表征便可以变得足够广泛且丰富,以至于能够应对在感知、想象和梦境中发生的意识表征。[13]
作为一种哲学立场,表征主义认为我们感知到的对象和特征并不局限于物理的外部世界,也不仅仅存在于大脑本身(例如,如果我们意识到橙色,那种颜色实际上并不存在于大脑中:神经元不会变成橙色)。相反,表征主义认为这些特征是我们在大脑外部体验到的意识表征的一部分,就如同这些特征被“投射”到我们周围的环境和我们的身体上一样。从这个立场出发,神经表征主义识别出那些作为意识现象生成的主要解释目标的神经机制。神经表征主义采用预测处理的计算原则来建立基于感觉的推理表征学习模型,并认为运动动作本身并不是意识所必需的[12],[13],[53]。尽管它与基于预测处理的主动推理意识解释有所区别,后者同样基于预测处理,但认为运动动作(或“能动性”)是意识的核心[10],[110]。在下面,我们将重点讨论神经表征主义或基于预测处理的解释。
广泛的证据表明,预测或表征是实现许多感觉、运动和认知过程的关键大脑功能,其中一些过程是在有意识的状态下进行的,而有些则不是[12],[110],[111]。为什么有些表征有助于意识的形成而有些则没有?在神经表征主义中,意识现象具有五个不可或缺的特征,这些特征共同定义了作为一个多模态、情境化的、对智能体世界(包括其身体)调查的意识体验[12],[13],[53]。这种调查或“概览”(以多模态的方式)服务于但不等同于复杂的决策和目标导向行为的计划。多种模态通过多感觉整合以及模态分离共同贡献了这一空间性的概览,使得模态和子模态被体验为质的不同。
在神经层面上,这种概览是由多个感觉-皮质层级(躯体感觉、听觉、视觉等)介导的,这些层级各自具有前馈/反馈架构,并且彼此相互作用,并与运动和计划系统相互作用[13](图4)。在这个非常大的系统中,不仅单一模态内的前馈和递归处理的相互作用是预测处理所必需的,而且模态之间以及在移动主体中运动系统的长程和横向相互作用也是必需[112]。实际上,视觉皮层对感知刺激的循环反应波的主要部分可以解释为与运动相关的活动(例如, Allen等[113]和Oude Lohuis等[114],[115])。这种“超级推理”的配置在其他能够生成预测的大脑结构中是不存在的(例如,小脑);因此,这些结构不涉及意识。神经表征主义考虑了几种深度预测处理的神经实现,从严格的层级系统到异构层级系统[116-118]。在这一观点下,意识和注意是不同的过程:虽然多模态拓扑结构提供了基本的意识体验,但注意系统则在整体拓扑结构中突出了有限的处理流。
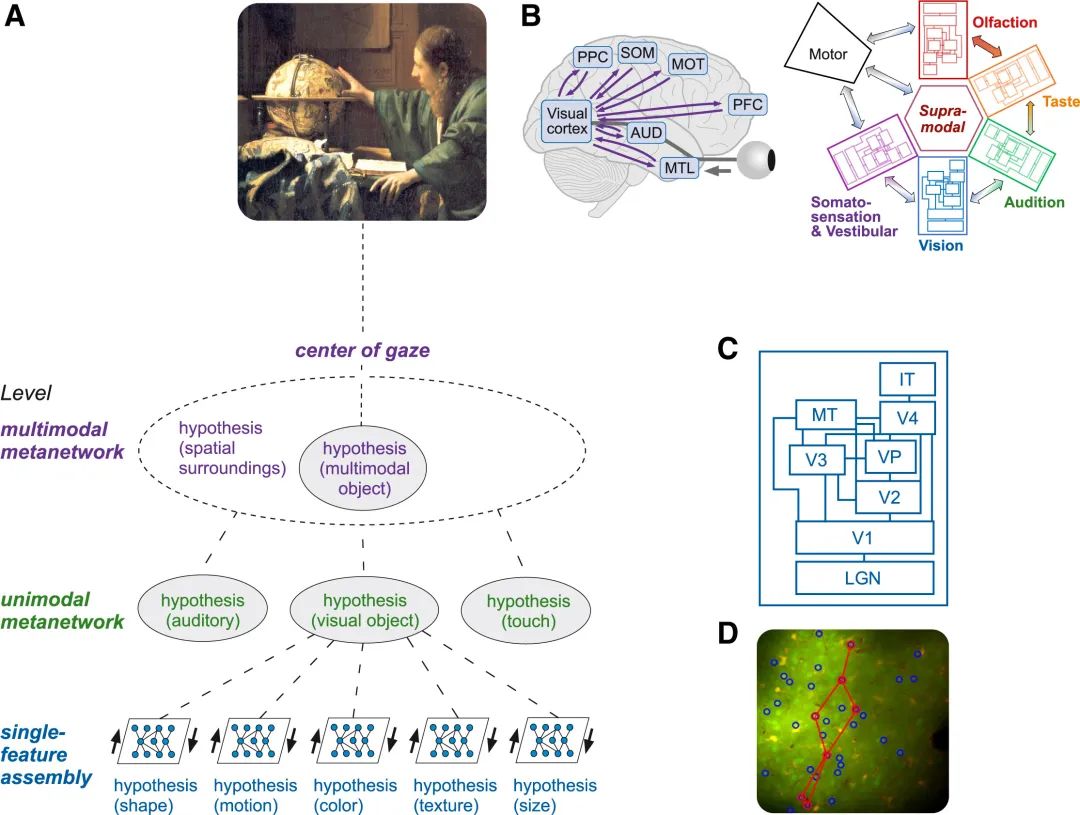
图4.与预测处理相关的神经表征主义(NREP)。NREP 认为,意识体验产生于多个表征层面,以维米尔(Vermeer)的画作《天文学家》(The Astronomer)的片段为例。
(A) 在底部,我们发现的是单一特征组件,作为最基本的表征层级(D 中的示例;在小鼠视觉皮层中记录的组件;参见 Montijn et al. [119])。这些组件执行预测编码操作,从而产生奇异特征(在本例中是视觉的:形状、运动、颜色等)的假设(表示)。这些特征表示被组合成单峰元网络级别的假设;在视觉中,这对应于主要在视觉皮层层次结构(C)中编码的联合表示。在多模态元网络层面,来自不同模态的假设被组合起来,并实现了跨空间的集成。这个水平对应于非凡的体验。
(B) 左:大脑的宏观视图,描绘了视觉皮层在视觉感知的起源中如何与其他皮质区域进行通信(AUD:听觉皮层;MTL:内侧颞叶;PPC:后顶叶皮层;SOM:体感皮层;对于身体活跃的受试者:MOT:运动皮层;PFC,前额叶皮层,参与目标导向的行为和计划)。
(C) 右图:不同的感觉皮质层次结构如何相互关联,以及在身体活跃的受试者的情况下如何与运动和执行区域进行通信的示意图。图改编自 Pennartz[13]和Pennartz et al.[112]。
神经表征主义的一个显著特点在于,意识表征具有多层次的性质。这一特征为意识难题提供了新的方法论视角:所谓的关键——“层次”,不应仅从神经解剖学或总量上理解(意识可以在不同的基质中实现,甚至是人工基质)。相反,层次在现象学上呈现出概念上的不同级别,自底向上,从单一神经元开始,逐步上升至集群层级、单模态及多模态网络的表征(图4)。
树突整合理论(DIT)[14],[56],[120]来源于对大脑皮层中循环处理机制的研究。它主要关注皮层深层的锥体神经元及其树突中的非线性过程,这些过程使得锥体神经元在皮层间及丘脑-皮层信息流中扮演了汇聚点的角色。目前,它基于六个核心观察,这些观察主要源于对啮齿动物皮质下投射(端脑外,ET)的第五层(L5)锥体神经元的研究:
1. L5锥体神经元通过非线性顶树突过程将到达单独的树突区室的信息关联起来,导致爆发放电。[120-122]
2. 抑制这些锥体神经元中的顶树突过程(例如树突状钙尖峰[dendritic calcium spikes])会抑制有意识的感知。[123]
3. 抑制 L5 锥体神经元的皮层下靶点(特别是高阶丘脑)会抑制有意识的感知。[124]
4. 麻醉引起的意识丧失与这些神经元中顶树突与基树突的解耦相关。[125]
5. 抑制高阶丘脑会使顶树突与基树突隔室解耦。[125]
6. 神经调节状态控制 L5 锥体神经元中的树突耦合。[125]
综合来看,树突整合理论认为,对这些趋同发现的最佳解释是,向皮质下投射的锥体神经元的顶轴,是皮层间和丘脑-皮层信息流的支点,这两者同时对哺乳动物的意识至关重要。[14],[56],[57]因此,理解这些L5神经元树突中突触输入整合的多个过程,对于理解其输出为何与意识如此紧密相关,至关重要。
树突整合理论的基本见解是,皮质L5锥体神经元在两个不同区域——顶区和基区接收和处理截然不同的信息,这两个区域在物理上和功能上都是分离的。这两个区之间的电分离已经很强,通过高“泄漏”导电性,使得顶区与体细胞区的计算活动基本上彼此隔离。然而,现在我们已经很清楚,这些神经元可以通过依赖树突中电压敏感离子通道的高度非线性过程,将两个区接收到的信息关联起来[121],[122],并由非常具体的抑制和神经调节作用在树突树的亚区上来控制。[126]
树突整合理论首先假设,这些神经元的基区主要接收前馈信息,这些信息与处理L5锥体神经元柱中的特定认知或感官特征有关。其次,它提出,长距离反馈信息主要作用于相同神经元的顶区,提供关于大脑其他地方(包括皮层其他区域)正在发生的过程的信息(或上下文)。
树突整合理论的一个独特之处在于它解释了哺乳动物大脑中信息流所需的多区神经元,这导致了许多推论和预测。[14],[120],[127]例如,树突整合理论的一个推论是,先前的知识(例如语义记忆)与感知过程的关联应依赖于连接到顶区的突触的影响。最近在啮齿动物中获得的证据表明,巩固的语义记忆储存在第 1 层的连接中,并影响顶树突活动。[128],[129]此外,高阶丘脑-皮层投射维持顶树突和基树突之间耦合的观察,被提出用以解释皮层间和丘脑-皮层环路的同时完成,因此可以作为解释(麻醉引起的)的意识丧失的机制。[14]
这反过来引出了一个假设,即定向注意作为访问意识的一个方面,是通过选择性维持不同皮层柱中的树突耦合来控制的,这样可以调节与现象意识相关的后皮层区域中锥体神经元的耦合。抑制和神经调节输入对耦合的调节还对大脑状态和意识(包括做梦)之间的关系提出了具体的预测。[130]
图5.树突整合理论 (DIT)。DIT提出特定的细胞机制是意识和与意识处理相关的全局激活模式的基础。
(A)皮质锥体细胞整合来自顶端和基底树突区室的信息。在两个隔间之间,有一个连接点(连接点),其作用类似于门并控制它们之间的耦合。
(B)左图,这种耦合机制控制丘脑皮质系统内的信息流。在意识状态下,锥体细胞内的门是打开的,允许信号传播到高阶丘脑,丘脑本身投射到中皮质层,维持顶端树突之间的耦合。是的,在无意识状态下,皮质锥体细胞解耦,导致丘脑皮质系统的活动传播崩溃。
(C)因此,投射到高阶丘脑的深层锥体神经元的顶端树突代表了整个皮层反馈的维持和影响的连接点。麻醉剂对该连接点(顶端树突的耦合区)的破坏会导致整个系统的反馈广泛丢失。
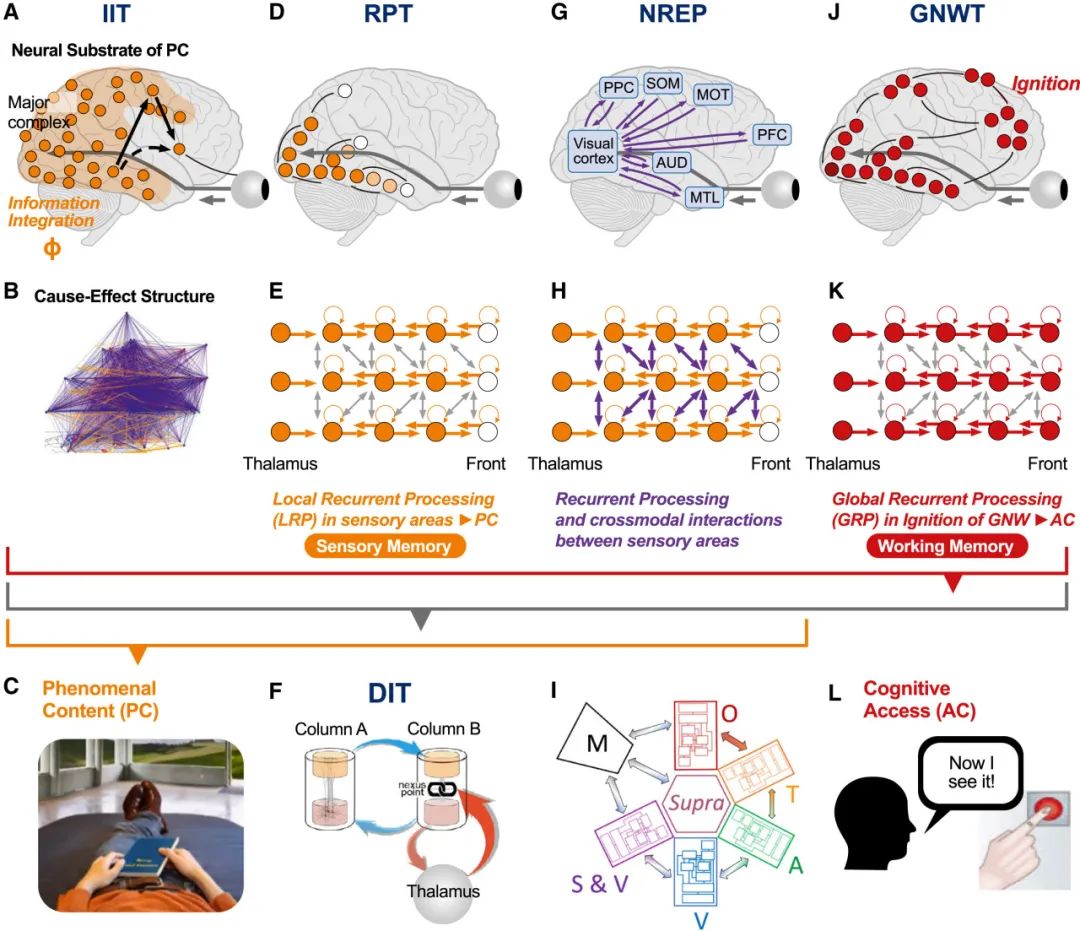
在这里,我们对所有五种理论进行了比较,讨论它们的相似性和差异性,以及它们的兼容性和互补性。所有理论都提出了自己的神经机制,这些机制通常显示出一定的重叠或相似之处,为理论间的融合提供了可能的结合点(有关差异,另见背景知识3)。神经机制可以在不同的组织层级上描述,从微观层次的(亚)细胞过程,到中观层次的区域内回路,再到宏观层次的大脑区域间相互作用。尽管不同的意识理论主要集中在不同的层级上,这些层级都被视为一个层次化的组织结构,使得描述它们的机制关系以及辨别差异和共性成为可能(见图 6)。
背景知识 3:如何推进意识理论:仍然存在的分歧、批评和方法
当前对神经意识理论的整合方法并不意味着我们可以对那些基本的差异和批评持一种天真的态度。除了对现象意识与访问意识的区别进行聚焦外,还有其他几个关键的差异值得关注。例如,全局神经工作空间理论(GNWT)、循环处理理论(RPT)、预测处理与神经表征主义(PP/NREP)以及树突整合理论(DIT)都强调信息处理、计算和消息传递在产生体验内容中的作用,而整合信息理论(IIT)则将体验与神经基质支持的因果结构联系起来。这需要考虑的不仅是基质的实际状态,还有其潜在状态的范围[95],而其他理论则将具体的意识体验解释为实际状态,而不考虑系统的所有可能状态。
因此,整合信息理论并不将意识识别为某种特定功能(见Tononi和Koch[131]),而其他理论则以功能性术语描述意识的神经基础,例如信息分布(GNWT)、学习和特征绑定(RPT)、预测(PP、DIT 和 NREP)、以及多模态情境调查(NREP)。然而,这些理论和其他理论将功能方面归因于意识,并不等同于计算功能主义(如Pennartz[12-13])。
对神经意识理论的一般批评是,在考虑了图1-7中描述的机制后,人们仍可能问道,“……但为什么这些机制会产生意识?”这个问题回到了“解释性鸿沟”或“难题”,各个理论以不同方式处理这个问题(见正文),但也应该认真对待这一批评。例如,普遍接受的再处理原则可能对意识是必要的,但不太可能是充分的。因此,当前的理论可能约束不足,需要基于实证证据和理论推理进一步完善。一个例子是,缺乏约束可能会导致非神经基质对意识的不同宽容度(例如,整合信息理论相对宽容,而神经表征主义和树突整合理论更为严格[132-133])。
此外,除了“结构”和“消息”之间的区分外,消息如何获得意义(意向性,“关于性”)的问题仍然悬而未决(如Pennartz[12],Chalmers [15]和Searle [134])。如何穿越这个看似令人生畏的哲学雷区?一种方法是对理论进行对抗性测试,测试它们在经验上可验证的预测点。Templeton 世界慈善基金会赞助了五个这样的项目,包括对GNWT、IIT、PP/NREP和 RPT的测试。实验目标包括代表意识内容的解剖位置、刺激持续时间和激活过程,[5]以及非活动神经元和运动活动在视觉空间感知中的作用(参见
https://www.templetonworldcharity.org/projects-database/0646)。
然而,我们认为这种对抗性测试将受益于补充方法,因为它可能(1)测试与被审查理论关系不大的预测,无法测试其核心假设,并且(2)过度依赖于证伪主义,期望对理论进行明确的拒绝/接受二分法。一个理论下被认为是错误的东西在另一个理论下可能是完全可以接受的,理论的改进可能更依赖于支持或反对某些解释的逐步证据,符合溯因推理(“对数据的最佳解释的推论” [135, 136]和贝叶斯证据积累[137])。因此,我们提倡一种补充方法,通过积累新的实证证据,将各种理论中最支持的元素结合起来,形成一个具有更强预测能力的改进框架。最后,我们还期望这些综合方法逐渐消除一些看似深刻的概念(有时是形而上学)鸿沟。例如,定义更多基于实证的理论约束,预计会限制能够合理归因于意识的复杂自然系统的范围。
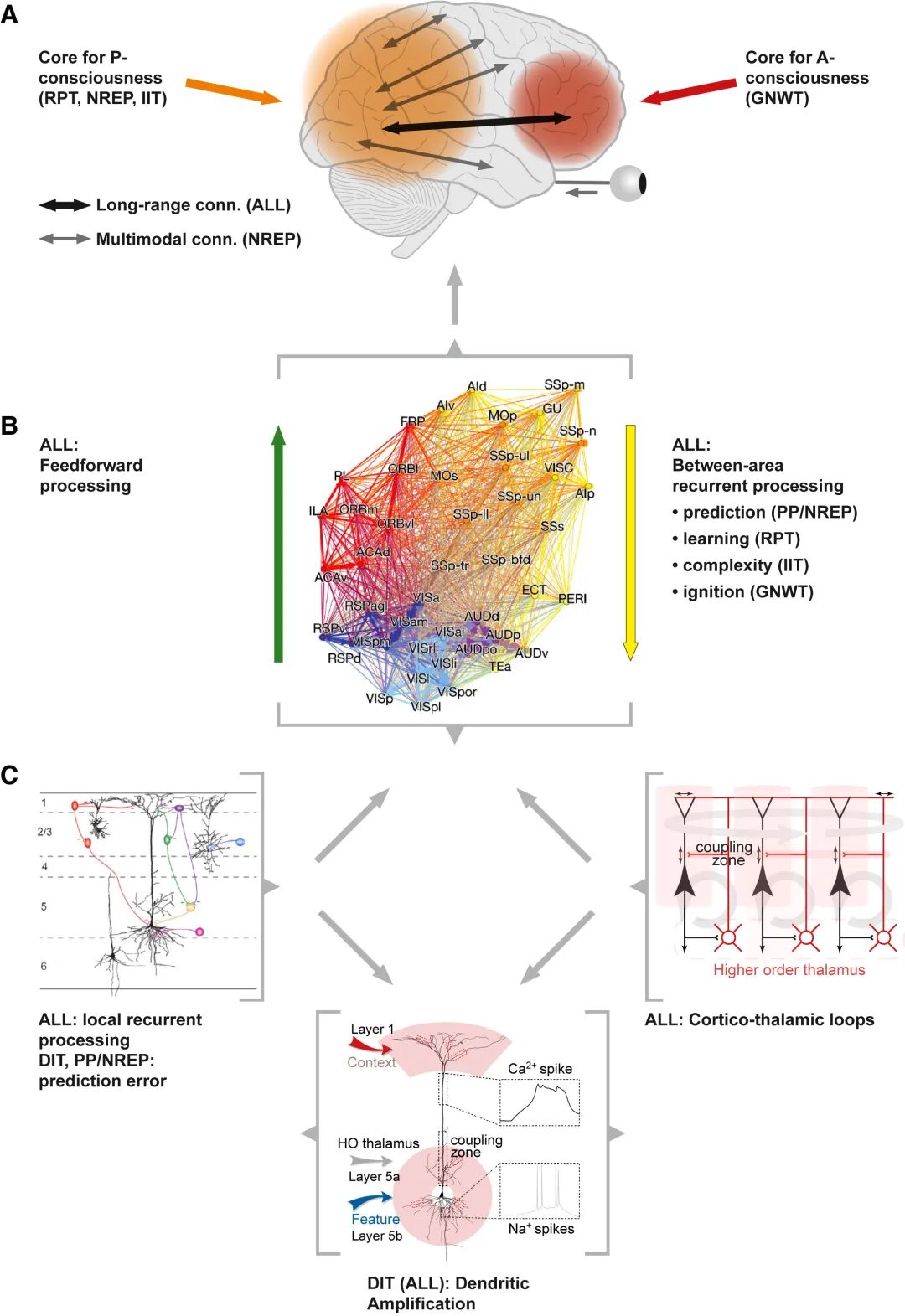
最低层(图C底部)由皮质锥体细胞代表,这些细胞中顶端树突放大的过程(如树突钙离子尖峰和树突-体细胞耦合)被树突整合理论(DIT)特别提出,为意识处理奠定了细胞基础。尽管其他四种理论未具体提出此基础,但“ALL”暗示这是一个共通点。下一个层级是皮质微电路(图C,左上[138])和皮质-丘脑环路(图C,HO:高阶)中的局部(区域内)循环处理。
(B)多区域水平由哺乳动物皮层[139]的大规模连接图表示,其中理论认为循环处理对于意识很重要,尽管各理论对此的解释不同。
(A)在宏观尺度上,橙色阴影区域表示涉及现象视觉体验的后皮质区域(根据循环处理理论、神经表征主义和整合信息理论;包括顶叶和颞叶区域),而全局神经工作空间理论提出前额叶区域,其中与更靠后区域的远程连接是访问意识的关键。神经表征主义通过灰色箭头假设了视觉区域与其他感觉模式和运动系统之间的基础通信。灰色括号勾勒出这些特定的总体层级;灰色箭头表示不同级别间的层次关系。
在诸多意识科学的共识中,有这样一种假设:意识最好在大规模网络及其计算层面上理解。然而,在大脑中,整体活动依赖于回路、细胞,甚至亚细胞层面的过程。在这里,我们采用多尺度方法[13],[140],试图从细胞水平开始揭示一些潜在的意识过程(参见图5-6)。
在神经系统的所有层面上,循环处理和整合是大多数意识理论的核心,包括树突整合理论、循环处理理论、全局神经工作空间理论、整合信息理论以及预测处理/神经表征主义。
特别是树突整合理论,它认为通过锥体神经元顶树突的非线性过程,循环处理和整合不可分割,构成了皮层-皮层和丘脑-皮层环路的关键枢纽(参见图6 C)。在清醒的、有意识的状态下,皮质L5锥体细胞处于“耦合状态”,使得到达顶树突的反馈可以影响细胞水平的体细胞处理,进而使反馈中的内部信息得以在整个系统中大规模并行地重新整合。[14],[125]根据树突整合理论,与意识状态相关的树突整合确保整个丘脑-皮层系统的活动能在空间与时间上得到整合。[56]这可能意味着活动可以通过皮层-皮层的循环连接传播和扩散,锥体细胞可以激活高阶丘脑核,这反过来又可以强化皮层中的活动。[14],[57]
根据树突整合理论,不断发展的活动产生了在宏观尺度上测量到的丰富的意识全局动态。[130]如果 L5 锥体细胞脱耦(例如,在麻醉或无梦睡眠期间),这种全局协调就不会发生;在这种情况下,活动可能仅在皮层内部传播,而不涉及更广泛的丘脑-皮层环路。[130]因此,与意识相关的大规模大脑动力学(通过fMRI、EEG测量)依赖于在亚细胞层面上发生的树突整合。
在树突整合理论中,这种局部机制为与其他处理层面相关的意识过程奠定了基础。[14]例如,整合信息理论预测,锥体细胞的脱耦会导致主复合体内因果效应的剧烈崩溃,降低Φ值,从而降低意识水平。[7]因此,树突整合为控制丘脑-皮层系统中信息整合提供了一个具体的细胞机制,这与计算建模一致。[54]
同样,如果树突整合对于丘脑-皮层网络中的信息流至关重要,那么如果额顶叶网络中神经元的顶端和基底部分脱耦,全局工作空间活动就会崩溃。[14],[125]因此,只有当L5锥体细胞处于耦合状态时,足够的活动传播和整合才能在丘脑-皮层系统中引发大规模激活。同样,局部树突整合可能使得在多个感觉模态和跨尺度上建立联合神经活动模式,这在神经表征主义中至关重要。总之,亚细胞树突整合可能是跨皮层或丘脑-皮层网络进行大规模整合的一个前提条件。因此,树突整合理论提出的机制可以被纳入任何依赖这种大规模整合的意识理论中。
树突整合还为具有双流架构的意识理论(如循环处理理论和经典预测处理;参见Rao和Ballard[141])提供了神经生物学细微差别。特别是关于树突整合机制的知识,具体说明了这两条流在单个锥体神经元水平上的相互作用。根据循环处理理论,意识来自于大脑中不同处理层次之间的反复相互作用。这一想法与树突整合理论兼容,因为锥体神经元的脱耦导致大脑中循环处理的崩溃。[14],[125]根据与预测处理相关的意识解释(如神经表征主义),从高到低处理层次发送的预测试图通过抑制匹配预测的感觉信号来“解释”从低到高处理层次传播的活动。不能预测的部分作为预测误差传递到更高层次。
虽然根据树突整合理论,L5锥体细胞固有的生物物理过程规定它们放大这两条流之间的匹配,[14],[120]但锥体神经元或邻近的皮层中间神经元的生物物理特性可能有多种方式被纳入预测处理框架(参见图6C[135],[142-144],[145])。所有这些变体都与树突整合作为生成意识体验的细胞基础的原则兼容。然而,根据树突整合理论,循环处理只是意识的一个必要条件。另一个关键组成部分是树突整合机制,它使得这些反馈信息能够融入丘脑-皮层系统的持续激活模式中。
皮质-丘脑微回路内外的局部处理,如何助益于意识的形成?除了树突整合理论强调的大型L5神经元外,在皮质柱内外,由数百万个突触连接的多种神经元类型形成了极其复杂的微回路,[146]这种复杂性很容易被低估。这种密集的复杂性,加上同样丰富的区域间和皮质-皮下的相互作用,可能超出人类的直觉,并且意外地为主观体验的看似非物质现象做出贡献。皮质神经元和微回路内外的局部、嵌套的复杂性对于本文讨论的所有五种理论都是相关的,例如,根据整合信息理论的说法,这种复杂性可能支持具有高因果效应(Φ)结构的现象意识。[95]
一般来说,无论是在微回路内[147]还是在更广泛的范围内,循环处理都是大多数意识理论(循环处理理论、全局神经工作空间理论、整合信息理论、预测处理/神经表征主义;参见“宏观层次”和“总体概念”一节)中的一个重要特征,并且也有助于与意识相关的过程,例如工作记忆(图 3)和感觉适应。[20]但具体来说,局部处理究竟能为意识贡献什么?基于“微观层次”一节中树突整合理论预测,预测处理/神经表征主义提出,低层次的预测和预测误差的计算,在很大程度上依赖于调节L5信息传输的局部柱内回路,但也包括L2/3锥体细胞。越来越多的证据表明,浅层锥体细胞在表示(预测)和误差编码中起作用,而L5细胞可能特别在表征编码中发挥作用。[23],[118],[135]特别是,生长抑素阳性中间神经元在误差计算中起到了作用。[148],[149]
虽然循环处理理论主要关注视觉区域内和之间的循环处理,[150-152]Lamme也提出,即使是网络中相互连接的神经元对之间的反馈处理也可能产生某些意识形态。然而,反馈的普遍性使Lamme提出,可能还需要其他过程,比如神经可塑性,来产生现象意识。[102]
全局神经工作空间理论表示,有意识的访问(AC)依赖于循环处理,因为它的“激活”和回响活动实现了对意识内容的访问。[28]该理论的早期版本提出,具有长程连接的L2/3锥体细胞(而不是树突整合理论中的L5神经元)对访问意识最为关键,但较新的版本也考虑了与皮质下结构的循环。Dehaene等人的访问意识模型,[153]包括具有层状前馈和反馈投射的丘脑-皮质柱,用P300波和注意瞬盲模拟了激活。
网络模型[154]提出了如何通过同一回路的不同操作模式访问和广播到工作记忆和意识(AC)的一个短暂的(<1 秒)感知缓冲区。根据整合信息理论,反馈/循环处理(RP)是意识所需的。根据计算Φ以“量化”现象意识的方法,具有纯粹前馈架构的人工系统的Φ=0,因此缺乏现象意识(PC),即使它们的外在行为(输入-输出功能)与具有反馈的有意识(Φ>0)系统不可区分[94-95](但参见Doerig[155])。
因此,如果反馈连接功能失效,即使在存在适当架构的情况下,意识也会消失。另一方面,由于整合信息理论认为脑回路的因果结构,而不是其活动,决定了Φ和现象意识,因此即使其神经元是静默的,主Φ复合体内因果整合的回路也可以对现象意识做出贡献。这种违反直觉的预测,没有其他理论所认同,正在整合信息理论和预测处理之间的对抗性合作中进行测试(见背景知识3)。
Haun和Tononi提出,感觉皮层中类似于晶格的“网格状”神经回路形成了空间体验的基质。[97]这种网格状连接在视觉和其他感觉区域中都有发现,但预测处理通常将感觉皮层的拓扑结构解释为从一个感觉空间到另一个进行推断的布局(参见Knierim和Van Essen[156])。此外,神经表征主义提出了主观空间体验源于在不同空间框架(视网膜坐标、头颅坐标和异中心[12])中设置的地图之间的多模态整合。
除了表示信息内容外,局部回路还随着不同“水平”的唤醒状态(反映了主要的“经典”意识“维度之一[157]”;参见Hill、Tononi[158] Laureys[159]和Bayne[160])经历神经调节状态的变化。然而,根据更近期的神经调节状态观点,这些可以被视为“背景条件”或启用因素,与意识本身只有间接关系(不具体说明意识内容)。[16]麻醉或神经调节引起的局部“唤醒”状态和网络复杂性的变化的影响已在丘脑-皮质网络模型中进行了模拟,[158]并且似乎与本文讨论的所有五种理论大致兼容,尽管尚未包括树突机制(DIT)。这种调制效应甚至在隔离的皮质切片(“中尺度[36]”)中通过改进版的扰动复杂性指数(PCI)和类似于在人类中使用的复杂性指数进行了评估(见背景知识1;“总体概念”一节)。尽管这些测量不能推断大脑切片和啮齿动物的意识,但它们可以提供有关负责大脑状态变化的细胞和网络机制的信息,这些变化伴随着复杂性的丧失和恢复。[161]
当我们逐层深入神经解剖的尺度,便会遇到这样一个问题:在脑区与由多个区域构成的系统层面上,支持意识体验的脑机制究竟是如何运作的?我们在此回顾的五种理论在这一层面上可分为三个部分。
首先,整合信息理论、预测处理/神经表征主义以及循环处理理论在处理意识视觉方面有共同之处,即它们均强调后皮质区域——特别是涉及视觉的枕叶、顶叶和颞叶——的关键作用。
整合信息理论认为,当高层和低层视觉区域之间发生密集的回返互动(reentrant interaction)时,最大Φ值将主要在后皮质(“热点区”)达到[78],[95]。基于预测处理的解释,包括神经表征主义,意识视觉始于视觉皮层层级内前馈信号流(主要传递错误信息)和回返信号流(传递自上而下预测)之间的互动,[23],[141],[135]尽管可能存在替代方案(例如Heeger[162])。整合信息理论和神经表征主义都认为,在一个可变的视觉环境中,不同的、可切换的脑区子集将参与前馈和反馈互动,以确保视觉特征的整合,这取决于被试体验中的视觉亚模态[12],[88]。在视觉皮层系统内的反馈处理也是循环处理理论达到现象意识的关键,同时需要全局回返处理以适应注意效应并达到访问意识。
其次,全局神经工作空间理论将意识视觉的突出作用归于前额和顶叶皮层,这与其对访问意识而非现象意识的关注,以及其将这种关注转化为行为反应、注意、工作记忆和评估等执行功能一致——所有这些功能都与前额-顶叶网络相关联[6]。
第三,如我们所见,树突整合理论更加侧重于意识的细胞基础。从这个意义上说,树突整合理论在关于意识视觉神经基质的“前部与后部”辩论中保持“中立”。值得注意的是,所有五种理论都同意,新皮层是产生意识内容的关键结构(与其他强调中枢结构的理论相对[163],[164],[165]),且丘脑核在维持意识方面是至关重要的。此外,所有五种理论都认为,除了局部或短程互动外,意识还需要依赖于皮层-丘脑系统中的长程通信[6],[7],[13],[105]。
然而,重要的是,循环处理理论和神经表征主义都认为,单凭视觉皮质层级内的信息处理,不足以实现有意识的视觉(无论是理解为现象意识还是访问意识)。根据循环处理理论,广泛的循环处理需要注意力的参与。而为此,视觉皮层和前额区域之间的互动被认为是必要的[102]。从这个意义上说,循环处理理论与全局神经工作空间理论在处理方式上的相似之处远超过它们定义上的不同:它们都强调调节注意信号的前额-顶叶网络的关键作用[102,105]。预测处理理论将注意解释为大预测误差和低决策信心的产物,促使被试在感觉空间中感知不确定的区域中寻求更多信息(例如Kanai等[166])。
在神经表征主义描述的意识体验特征中,我们看到了动态和稳定性的结合,其中“稳定性”是指,即使在眼、头和身体处于运动中,我们也能将外部世界及其物体感知为一个稳定的整体[12,13]。为了在身体活动的被试中实现视觉稳定,需要视觉和运动皮层之间的互动:视觉系统接收运动信号(以及本体感觉和前庭回馈)或关于哪些感觉变化正在发生或即将发生的预测[112,166]。此外,神经表征主义假设感觉皮层系统之间的横向、多感觉互动是确保意识的定性、多模态丰富性所必需的[167]。因此,视觉体验并非仅由传统定义的视觉皮层-丘脑系统介导,而是由包括许多卫星区域的“扩展视觉皮层”介导。整合信息理论和循环处理理论在其他方面并不与这些附加互动矛盾,但并未特别强调这一点。
因此,乍一看,这三种理论在意识视觉的宏观解剖学上至少总体一致,而另外两种理论则要么表达了不同的见解(全局神经工作空间理论与背景定义的差异有关),要么保持中立(树突整合理论)。这种整合信息理论、循环处理理论和神经表征主义的全局兼容性,并不意味着它们具有相同的范围或相似性。例如,它们使用不同的理由来解释为什么后皮质的回返反馈对于意识是必要的(整合信息理论是为了最大化Φ;循环处理理论是为了实现可塑性、学习和特征绑定;神经表征主义是为了闭合表示学习和推理的循环)。另一个关键区别在于对非活跃神经元的假定作用:(在整合信息理论中,除非它们与网络断开连接或失活,否则这些神经元可以对意识体验做出贡献[55];其他理论不遵循这一点)。尽管如此,我们可以得出结论,在多区域系统层面上,这些理论并不像有时认为的那样不相容。
越来越多的实验证据表明,与意识相关的神经元互动模式,不仅需要通过前馈和反馈连接紧密集成,还必须是差异化和信息丰富的。[32],[168],[169]这种大脑复杂性的概念——定义为集成和差异化的共存——是整合信息理论中原始概念Φ的核心[89],[90],并在其成熟形式的经验预测中继续占据中心位置。[95]
最近,这一复杂性的概念亦被其他理论所接纳。例如,全局神经工作空间理论的一项关键原则便是广泛的神经激活。因此,整合信息理论与全局神经工作空间理论实际上在神经元集成的基本需求上达成共识,尽管两者的理论基础存在差异。值得注意的是,全局神经工作空间理论的初版并未明确提及与信息相关的度量、差异化或复杂性,其主要关注的是神经过程,如额-顶叶的同步及P3b波。然而,随后全局神经工作空间理论的支持者的研究将重点转移到了动态系统的丰富性[30],[170],并且最近,复杂性的概念被明确纳入体系,视其为基于分布式模块广播不同类型感觉信息的关键元素。[171]
根据整合信息理论的定义,大脑复杂性与树突整合理论和循环处理理论所假设的微观和中观层级具有明确的机械关系。这一过程需要有效的回返互动,确保差异化的神经元群体紧密结合并参与丰富的因果互动。树突解耦和循环互动的破坏预计会导致大脑集成信息能力的急剧下降。在这方面,睡眠期间和无反应觉醒综合征(“植物人”状态)患者中观察到的复杂性崩溃与反馈互动的破坏有关。[37],[172-174]
类似地,复杂性是神经表征主义假设的多模式丰富性和集成的内在特征,尽管也有重要的差异。在整合信息理论中,大脑复杂性解释了每一种体验中“即时”的普遍现象特性(统一性和丰富性),而神经表征主义和预测处理一般将有意识的体验视为对外部现实的代表性“模拟”。[12],[13],[175],[176]在神经表征的观点中,复杂性转化为一个多方面的现象,包括有意识内容的多模式、定性丰富性、空间延展性和分辨率、强度和注意力,以及最终的时间深度——所有这些都被理解为有利于后续的行动计划。[13],[53]在这方面,有很多方法可以定义和操作化大脑复杂性,更多的努力是为了指定基础假设并将复杂性的概念扩展到一个详尽的分类法中。[32]
尽管如此,关于复杂性这一概念的整体趋同,[88]近来显示出了一些引人入胜的意义——特别是在无反应患者的经验应用上[170],[177],[178](背景知识1)。广泛的理论一致性可能会增加这种信心,也在更具挑战性的情形中证明了其推理的合理性。此外,复杂性与信息丰富性的研究可在多个层面展开(图 6),从人类颅外和颅内记录到啮齿动物[38],[169],[179-181]和皮层切片。[182]在人类大脑之外,当前的复杂性度量无法对意识水平进行任何推论。然而,它们为在不同尺度上建立神经过程之间的机械联系提供了可能,从微观的树突整合到中观的递归过程,再到宏观的区域间集成。最终,这种关于复杂性的趋同或许代表了一种比较不同框架的共同基础。
在大脑中,复杂性何处体现,又是如何实现的呢?对于整合信息理论来说,具有循环金字塔状、收敛-发散连接(如在后部区域发现的那样)的网格状皮层的因果结构,被认为对应于复杂性的最大值,因此足以实现有意识的体验。[96],[97]预测处理-神经表征主义与整合信息理论共同追求空间延展性感知的神经基础,但它们在枕叶、颞叶和顶叶皮层所使用的视网膜定位、头颅定位和以物为中心的参考框架之间寻求整合;这些不同的方向可能被证明是兼容的。
另一个关键问题是:为什么有些意识理论关注持续的神经元活动的复杂性,而另一些则强调底层因果结构的复杂性?[183]澄清这一方面很重要,因为它不仅涉及不同的测量方法(观察性vs.扰动性),还涉及对意识中重要信息类型的实质性不同理解(外在信息vs.内在因果结构)[7,184,185](背景知识3)。最后,当前理论如何考虑分析大脑内因果互动的结构和复杂性,也可能有助于寻找内容特定的意识神经关联?尽管这一问题具有挑战性,但已经在这个方向上进行了尝试。[97],[167],[176]
图7.比较意识理论。本图说明了本文讨论的五种意识理论如何部分互补、部分重叠,以及它们如何关联核心概念和解释。这些理论主要围绕现象意识(PC;橙色元素和C)和访问意识(AC;红色)元素和L),这里主要用于视觉说明。
A和B:整合信息论(IIT)及其综合信息测量(Φ),这是根据 IIT量化现象意识(P意识)的方法。C:用一只眼睛的主观视觉体验的图像来说明现象意识(PC)[186]。D 和 E:循环处理理论(RPT)。F:树突整合理论 (DIT)。G、H 和 I:神经表征主义(NREP);其中I中的缩写可参见图4B的解释。J 和 K:全局神经元工作空间理论(GNWT)。L:通过对感官刺激的行为和认知反应来说明访问意识 (AC)。
我们认为这五种理论在解释意识的不同方面时是部分互补的:IIT、RPT 和 NREP 主要致力于解释现象意识;而全局部神经元工作空间理论主要关注解释访问意识(,这也是其他理论的关注点。(B、E、H 和 K 下的红色括号)。
F:树突整合理论被提议为大脑网络功能提供细胞基础,这些功能似乎是意识所必需的(PC 和 AC),并且与所有其他理论兼容:IIT、RPT、NREP 和 GNWT(B,E,H和K下的灰色括号)。E、H和K中带箭头的彩色圆圈水平阵列代表前馈和循环处理流;中间行代表从丘脑(LGN)到额叶皮层(前)的中央视觉通路;上行和下行代表辅助视觉通路(RPT、GNWT和NREP)或其他感觉模式(NREP)。RPT认为视觉系统内的循环处理及其高级区域是视觉现象意识的核心。RPT和GNWT认为,循环处理也必须涉及前额叶系统才能达到访问意识。NREP强调感觉模态和子模态之间的串扰来解释现象意识的定性差异,即使这涉及意识视觉(H,紫色箭头)。
尽管表面上看来,调和众多意识理论,甚至形成一个统一的意识理论似乎是不可能的挑战——至少有些理论表现出的差异如此之大(参见背景知识3),以至于弥合它们之间的鸿沟似乎是难以逾越的。这甚至导致了竞争理论之间的不合法性指控。[187]但是,我们的立场是,这些理论的多样性反映了这一新兴领域的健康发展。密切关注被解释的现象可以解开核心主张,使多个理论能够共同为我们的理解做出贡献。理论的重点和解释现象的基本差异有时可能使它们更加兼容。例如,全局神经工作空间理论专注于访问意识,而循环处理理论、整合信息理论和预测处理-神经表征主义主要旨在解释现象意识。
一个综合的意识理论需要以神经的术语解释现象意识,同时也要解释意识内容如何分配并用于后续的行动计划、工作记忆、价值评估和注意力调节等,这是访问意识的领域。目前研究者确实在这方面进行了一些建模工作,例如将预测处理的解释与全局神经工作空间理论的操作结合起来。[188],[189]我们还看到,在原则上,循环处理理论可以与全局神经工作空间理论整合,共同将访问意识纳入其中。由于整合信息理论已经承认前额叶区域在意识访问中的作用,因此在其“前部”关注访问意识方面与全局神经工作空间理论并不矛盾。尽管如此,有些人会认为全局神经工作空间理论与整合信息理论不能同时正确,因为它们在意识的本质上存在分歧(另见背景知识3)。
另一个潜在的概念汇合点在于高层次的、与复杂性相关的概念,如信息整合(整合信息理论)、全局信息可用性(全局神经工作空间理论)、多级表示中的多模态丰富性(预测处理-神经表征主义)、在不同解剖尺度上的循环处理(循环处理理论)和皮质-丘脑环路中的整合(树突整合理论)。虽然从表面上看,这些想法显著不同,但它们都从根本上基于相互关联性的不同方面,认识到依赖于皮质-丘脑系统中的短程和长程连接的神经元互动的丰富性,是显著程度的意识的重要因素(参见图7)。
这些汇合点和相关的考虑提供了某种希望,即一个综合的意识理论——或者至少一组相关的交织概念和工作假设——可能在未来几年或几十年内变得触手可及。我们也必须承认,大量的理论与实证研究领域仍待开拓(参见背景知识3)。意识到我们仅是初探这一巨大议题的表层,这很让人感到羞愧,但现如今我们显然已在多个方面取得了持续的进展。
当前的意识研究,犹如从不同角度审视一个多面体,每个角度都基于个人的视角强调不同的方面。此情此景,似可与量子力学相互类比,在那里对观察和现象有一致意见,但在如何解释概率的微观描述和我们确定性的宏观日常生活之间的关系上仍有广泛分歧。这两种情况可能都反映了人类想象力和直觉的根本限制。[12],[190]这种类比强调了意识研究者开发明确概念和共同术语的迫切需要(如将通用术语“意识”细分为“访问意识”与“现象意识”),并设计出将不同理论表述相互转化的方法。正如量子理论受益于坚实的数学框架,意识研究领域亦应如此。
最后,我们主张广泛且无争议地支持更多的实验测试与意识的实证测量的发展,这一点具有双重重要性。首先,如扰动复杂性指数(PCI)等新的、基于理论的实证测量,以及其他与复杂性相关的测量,能够有效应用于临床环境中,例如帮助对意识障碍的无反应患者进行更细致的分类,并区分他们不同的恢复与治疗前景(如Demertzi[170]Casarotto[177]和Farisco[191])。其次,实证测量有助于区分或统一不同的理论。在有意识状态下由经颅磁刺激引发的高复杂度时空电脑脑电图模式始终与循环激活波的存在相关,[174],[192]而在慢波睡眠和麻醉特征的皮层上升和下降状态下则发现了低复杂度模式(如Steriade[161]和Olcese[193])。因此,PCI不仅与整合信息理论的假设相符,原则上也与全局神经工作空间理论、循环处理理论、神经表征主义和树突整合理论相关的神经过程相兼容。[13-14],[102],[171]
该领域已经受益于更广泛的实证工具和测量方法。例如,树突整合理论引入了脑电图与局部场电位标记,与不同意识阶段的树突钙离子峰值对齐。[194]预测处理/神经表征主义提出了意识表示的测量,依赖于解码主体体验的有意识内容的技术(参见Goltstein[195];另见背景知识2),而全局神经工作空间理论有望从报告与无报告范式中受益,结合单细胞记录、功能性磁共振成像和脑电图-脑磁图(Sergent[42])。利用补充的计算方法可以进一步促进比较和整合,[116],[188],[196],[197]并且研究正在从人类扩展到动物意识,以受益于体内双光子成像、大规模群体记录、复杂性测量和光遗传学,揭示意识的因果基质(如van Vugt[64],Oude Lohuis[114-115], Takahashi[124],Suzuki&Larkum[125],Arena[38],[179],O’Connor[198]和Storm[199])。对抗性研究正在成为意识研究的重要新方法,[5]但目前尚不清楚这是否最终会导致单一获胜理论的达尔文选择(参见背景知识3)。在这里,我们提倡一种更加统一的、整合导向的方法,寻求通过实证测试和结合各种理论的有价值元素来推进研究。
总之,我们认为神经意识理论(1)显示出的差异往往不像最初想象的那样大或不可逾越,(2)提供了在不同组织层次上结合和整合各种元素的机会,以及(3)一个理解大脑-意识关系的综合框架,在实证和理论进展的指导下,具有在未来十年推动该领域发展的巨大潜力。
1. Chalmers, D.J. (1995). Facing up to the problem of consciousness. J. Conscious. Stud. 2, 200–219.
2. Crick, F. (1995). The Astonishing Hypothesis: the Scientific Search for the Soul (Simon and Schuster).
3. Crick, F., and Koch, C. (1990). Towards a neurobiological theory of consciousness. Semin. Neurosci. 2, 263–275.
4. Seth, A.K., and Bayne, T. (2022). Theories of consciousness. Nat. Rev. Neurosci. 23, 439–452.
5. Cogitate Consortium, Ferrante, O., Gorska-Klimowska, U., Henin, S., Hirschhorn, R., Khalaf, A., Lepauvre, A., Liu, L., Richter, D., Vidal, Y., et al. (2023). An adversarial collaboration to critically evaluate theories of consciousness. Preprint at bioRxiv. https://doi.org/10.1101/2023.06.23.546249.
6. Dehaene, S., and Changeux, J.P. (2011). Experimental and theoretical approaches to conscious processing. Neuron 70, 200–227.
7. Tononi, G., Boly, M., Massimini, M., and Koch, C. (2016). Integrated information theory: from consciousness to its physical substrate. Nat. Rev. Neurosci. 17, 450–461.
8. Lamme, V.A.F. (2006). Towards a true neural stance on consciousness. Trends Cogn. Sci. 10, 494–501.
9. Clark, A. (2013). Whatever next? Predictive brains, situated agents, and the future of cognitive science. Behav. Brain Sci. 36, 181–204.
10. Friston, K.J., Wiese, W., and Hobson, J.A. (2020). Sentience and the Origins of Consciousness: From Cartesian Duality to Markovian Monism.
11. Hohwy, J., and Seth, A. (2020). Predictive processing as a systematic basis for identifying the neural correlates of consciousness. PhiMiSci. 1, 1.
12. Pennartz, C.M.A. (2015). The Brain’s Representational Power: on Consciousness and the Integration of Modalities (MIT Press).
13. Pennartz, C.M.A. (2022). What is neurorepresentationalism? From neural
activity and predictive processing to multi-level representations and consciousness. Behav. Brain Res. 432, 113969.
14. Aru, J., Suzuki, M., and Larkum, M.E. (2020). Cellular Mechanisms of
Conscious Processing. Trends Cogn. Sci. 24, 814–825.
15. Chalmers, D.J. (2010). The Character of Consciousness (Oxford University Press).
16. Koch, C., Massimini, M., Boly, M., and Tononi, G. (2016). Neural correlates of consciousness: progress and problems. Nat. Rev. Neurosci.
17. Sanders, R.D., Tononi, G., Laureys, S., and Sleigh, J.W. (2012). Unresponsiveness s Unconsciousness. Anesthesiology 116, 946–959.
18. Nagel, T. (1974). What Is It Like to Be a Bat? Philos. Rev. 83, 435–450.
19. Block, N. (2005). Two neural correlates of consciousness. Trends Cogn.
20. Wang, X.J. (2001). Synaptic reverberation underlying mnemonic persistent activity. Trends Neurosci. 24, 455–463.
21. Hempel, C.G., and Oppenheim, P. (1948). Studies in the Logic of Explanation. Philos. Sci. 15, 135–175.
22. Carlson, N.R. (2010). Psychology: the Science of Behavior, Seventh Edition (Allyn & Bacon).
23. Bastos, A.M., Usrey, W.M., Adams, R.A., Mangun, G.R., Fries, P., and
Friston, K.J. (2012). Canonical Microcircuits for Predictive Coding.
24. National Academy of Sciences (2008). Science, Evolution, and Creationism (Washington, DC: The National Academies Press).
25. Klink, P.C., Self, M.W., Lamme, V.A.F., and Roelfsema, P.R. (2015). Theories and Methods in the Scientific Study of Consciousness (John Benjamins Publishing Company).
26. Koch, C., and Tsuchiya, N. (2007). Attention and consciousness: two
distinct brain processes. Trends Cogn. Sci. 11, 16–22.
27. Lamme, V.A.F. (2020). Visual Functions Generating Conscious Seeing.
28. Mashour, G.A., Roelfsema, P., Changeux, J.P., and Dehaene, S. (2020).
Conscious Processing and the Global Neuronal Workspace Hypothesis.
29. Bekinschtein, T.A., Dehaene, S., Rohaut, B., Tadel, F., Cohen, L., and
Naccache, L. (2009). Neural signature of the conscious processing of
auditory regularities. Proc. Natl. Acad. Sci. USA 106, 1672–1677.
30. King, J.R., Faugeras, F., Gramfort, A., Schurger, A., El Karoui, I., Sitt, J.D.,
Rohaut, B., Wacongne, C., Labyt, E., Bekinschtein, T., et al. (2013). Single-trial decoding of auditory novelty responses facilitates the detection
of residual consciousness. NeuroImage 83, 726–738.
31. Faugeras, F., Rohaut, B., Weiss, N., Bekinschtein, T., Galanaud, D., Puybasset, L., Bolgert, F., Sergent, C., Cohen, L., Dehaene, S., et al. (2012).
Event related potentials elicited by violations of auditory regularities in
patients with impaired consciousness. Neuropsychologia 50, 403–418.
32. Sarasso, S., Casali, A.G., Casarotto, S., Rosanova, M., Sinigaglia, C., and
Massimini, M. (2021). Consciousness and complexity: a consilience of
evidence. Neuroscience of Consciousness 2021, niab023.
33. Demertzi, A., Soddu, A., and Laureys, S. (2013). Consciousness supporting networks. Curr. Opin. Neurobiol. 23, 239–244.
34. Luppi, A.I., Craig, M.M., Pappas, I., Finoia, P., Williams, G.B., Allanson,
J., Pickard, J.D., Owen, A.M., Naci, L., Menon, D.K., et al. (2019). Consciousness-specific dynamic interactions of brain integration and functional diversity. Nat. Commun. 10, 4616.
35. Casali, A.G., Gosseries, O., Rosanova, M., Boly, M., Sarasso, S., Casali,
K.R., Casarotto, S., Bruno, M.A., Laureys, S., Tononi, G., et al. (2013). A
Theoretically Based Index of Consciousness Independent of Sensory
Processing and Behavior. Sci. Transl. Med. 5, 198ra105.
36. D’Andola, M., Giulioni, M., Dante, V., Del Giudice, P., and Sanchez-Vives,
M.V. (2019). Control of cortical oscillatory frequency by a closed-loop
system. J. Neuroeng. Rehabil. 16, 7.
37. Rosanova, M., Fecchio, M., Casarotto, S., Sarasso, S., Casali, A.G., Pigorini, A., Comanducci, A., Seregni, F., Devalle, G., Citerio, G., et al.
(2018). Sleep-like cortical OFF-periods disrupt causality and complexity
in the brain of unresponsive wakefulness syndrome patients. Nat. Commun. 9, 4427.
38. Arena, A., Comolatti, R., Thon, S., Casali, A.G., and Storm, J.F. (2021).
General Anesthesia Disrupts Complex Cortical Dynamics in Response
to Intracranial Electrical Stimulation in Rats. eNeuro 8. ENEURO.0343-
39. Rosanova, M., Gosseries, O., Casarotto, S., Boly, M., Casali, A.G.,
Bruno, M.A., Mariotti, M., Boveroux, P., Tononi, G., Laureys, S., et al.
(2012). Recovery of cortical effective connectivity and recovery of consciousness in vegetative patients. Brain 135, 1308–1320.
40. Aru, J., Bachmann, T., Singer, W., and Melloni, L. (2012). Distilling the
neural correlates of consciousness. Neurosci. Biobehav. Rev. 36,
41. Avanzini, P., Abdollahi, R.O., Sartori, I., Caruana, F., Pelliccia, V., Casaceli, G., Mai, R., Lo Russo, G., Rizzolatti, G., and Orban, G.A. (2016).
Four-dimensional maps of the human somatosensory system. Proc.
Natl. Acad. Sci. USA 113, E1936–E1943.
42. Sergent, C., Corazzol, M., Labouret, G., Stockart, F., Wexler, M., King,
J.R., Meyniel, F., and Pressnitzer, D. (2021). Bifurcation in brain dynamics
reveals a signature of conscious processing independent of report. Nat.
43. Dembski, C., Koch, C., and Pitts, M. (2021). Perceptual awareness negativity: a physiological correlate of sensory consciousness. Trends Cogn.
44. Avanzini, P., Pelliccia, V., Lo Russo, G., Orban, G.A., and Rizzolatti, G.
(2018). Multiple time courses of somatosensory responses in human cortex. NeuroImage 169, 212–226.
45. Del Vecchio, M., Caruana, F., Sartori, I., Pelliccia, V., Lo Russo, G., Rizzolatti, G., and Avanzini, P. (2019). Ipsilateral somatosensory responses
in humans: the tonic activity of SII and posterior insular cortex. Brain
Struct. Funct. 224, 9–18.
46. Del Vecchio, M., Fossataro, C., Zauli, F.M., Sartori, I., Pigorini, A., d’Orio,
P., Abarrategui, B., Russo, S., Mikulan, E.P., Caruana, F., et al. (2021).
Tonic somatosensory responses and deficits of tactile awareness
converge in the parietal operculum. Brain 144, 3779–3787.
47. Fossataro, C., Bruno, V., Bosso, E., Chiotti, V., Gindri, P., Farne` , A., and
Garbarini, F. (2020). The sense of body-ownership gates cross-modal
improvement of tactile extinction in brain-damaged patients. Cortex
48. Schneider, M., Kemper, V.G., Emmerling, T.C., De Martino, F., and Goebel, R. (2019). Columnar clusters in the human motion complex reflect
consciously perceived motion axis. Proc. Natl. Acad. Sci. USA 116,
49. Salzman, C.D., and Newsome, W.T. (1994). Neural Mechanisms for
Forming a Perceptual Decision. Science 264, 231–237.
50. Ellia, F., Hendren, J., Grasso, M., Kozma, C., Mindt, G., P Lang, J., M
Haun, A., Albantakis, L., Boly, M., and Tononi, G. (2021). Consciousness
and the fallacy of misplaced objectivity. Neurosci. Conscious. 2021,
51. Block, N. (2023). The Border Between Seeing and Thinking (Oxford University Press).
52. Sevenius Nilsen, A.S., Juel, B.E., Thurer, B., Aamodt, A., and Storm, J.F.
(2022). Are we really unconscious in ‘‘unconscious’’ states? Common assumptions revisited. Front. Hum. Neurosci. 16, 987051.
53. Pennartz, C.M.A. (2018). Consciousness, Representation, Action: The
Importance of Being Goal-Directed. Trends Cogn. Sci. 22, 137–153.
54. Munn, B.R., Muller, E.J., Aru, J., Whyte, C.J., Gidon, A., Larkum, M.E., €
and Shine, J.M. (2023). A Thalamocortical Substrate for Integrated Information via Critical Synchronous Bursting. Proc Natl Acad Sci USA 120.
55. Tononi, G. (2004). An information integration theory of consciousness.
56. Bachmann, T., Suzuki, M., and Aru, J. (2020). Dendritic integration theory: A thalamo-cortical theory of state and content of consciousness.
57. Aru, J., Suzuki, M., Rutiku, R., Larkum, M.E., and Bachmann, T. (2019).
Coupling the State and Contents of Consciousness. Front. Syst. Neurosci. 13, 43.
58. Baars, B.J. (1988). A Cognitive Theory of Consciousness (Cambridge
59. Dehaene, S., Kerszberg, M., and Changeux, J.P. (1998). A neuronal model of a global workspace in effortful cognitive tasks. Proc. Natl. Acad. Sci. USA 95, 14529–14534.
60. Dehaene, S., and Naccache, L. (2001). Towards a cognitive neuroscience of consciousness: basic evidence and a workspace framework. Cognition 79, 1–37.
61. Felleman, D.J., and van Essen, D.C. (1991). Distributed hierarchical processing in the primate cerebral cortex. Cereb. Cortex 1, 1–47.
62. Grill-Spector, K., and Malach, R. (2004). The human visual cortex. Annu. Rev. Neurosci. 27, 649–677.
63. Mesulam, M.M. (1999). Spatial attention and neglect: parietal, frontal and cingulate contributions to the mental representation and attentional targeting of salient extrapersonal events. Philos. Trans. R. Soc. Lond. B Biol. Sci. 354, 1325–1346.
64. van Vugt, B., Dagnino, B., Vartak, D., Safaai, H., Panzeri, S., Dehaene, S., and Roelfsema, P.R. (2018). The threshold for conscious report: Signal loss and response bias in visual and frontal cortex. Science 360, 537–542.
65. Noel, J.P., Ishizawa, Y., Patel, S.R., Eskandar, E.N., and Wallace, M.T. (2019). Leveraging Nonhuman Primate Multisensory Neurons and Circuits in Assessing Consciousness Theory. J. Neurosci. 39, 7485–7500.
66. Sanchez, G., Hartmann, T., Fusca` , M., Demarchi, G., and Weisz, N. (2020). Decoding across sensory modalities reveals common supramodal signatures of conscious perception. Proc. Natl. Acad. Sci. USA 117, 7437–7446.
67. Marti, S., King, J.R., and Dehaene, S. (2015). Time-Resolved Decoding of Two Processing Chains during Dual-Task Interference. Neuron 88, 1297–1307.
68. Marti, S., and Dehaene, S. (2017). Discrete and continuous mechanisms of temporal selection in rapid visual streams. Nat. Commun. 8, 1955.
69. Sergent, C., Baillet, S., and Dehaene, S. (2005). Timing of the brain events underlying access to consciousness during the attentional blink. Nat. Neurosci. 8, 1391–1400.
70. Berkovitch, L., Dehaene, S., and Gaillard, R. (2017). Disruption of Conscious Access in Schizophrenia. Trends Cogn. Sci. 21, 878–892.
71. Charles, L., King, J.R., and Dehaene, S. (2014). Decoding the Dynamics of Action, Intention, and Error Detection for Conscious and Subliminal Stimuli. J. Neurosci. 34, 1158–1170.
72. Del Cul, A.D., Baillet, S., and Dehaene, S. (2007). Brain Dynamics Underlying the Nonlinear Threshold for Access to Consciousness. PLoS Biol. 5, e260.
73. Marti, S., Sigman, M., and Dehaene, S. (2012). A shared cortical bottleneck underlying Attentional Blink and Psychological Refractory Period. NeuroImage 59, 2883–2898.
74. Gelbard-Sagiv, H., Mukamel, R., Harel, M., Malach, R., and Fried, I. (2008). Internally Generated Reactivation of Single Neurons in Human Hippocampus During Free Recall. Science 322, 96–101.
75. Kreiman, G., Fried, I., and Koch, C. (2002). Single-neuron correlates of subjective vision in the human medial temporal lobe. Proc. Natl. Acad. Sci. USA 99, 8378–8383.
76. Quiroga, R.Q., Mukamel, R., Isham, E.A., Malach, R., and Fried, I. (2008). Human single-neuron responses at the threshold of conscious recognition. Proc. Natl. Acad. Sci. USA 105, 3599–3604.
77. Reber, T.P., Faber, J., Niediek, J., Bostro¨ m, J., Elger, C.E., and Mormann, F. (2017). Single-Neuron Correlates of Conscious Perception in the Human Medial Temporal Lobe. Curr. Biol. 27, 2991–2998.e2.
78. Boly, M., Massimini, M., Tsuchiya, N., Postle, B.R., Koch, C., and Tononi, G. (2017). Are the Neural Correlates of Consciousness in the Front or in the Back of the Cerebral Cortex? Clinical and Neuroimaging Evidence. J. Neurosci. 37, 9603–9613.
79. Bellet, J., Gay, M., Dwarakanath, A., Jarraya, B., van Kerkoerle, T., Dehaene, S., and Panagiotaropoulos, T.I. (2022). Decoding rapidly presented visual stimuli from prefrontal ensembles without report nor post-perceptual processing. Neurosci. Conscious. 2022, niac005.
80. Kapoor, V., Dwarakanath, A., Safavi, S., Werner, J., Besserve, M., Panagiotaropoulos, T.I., and Logothetis, N.K. (2022). Decoding internally generated transitions of conscious contents in the prefrontal cortex without subjective reports. Nat. Commun. 13, 1535.
81. Kok, P., Bains, L.J., van Mourik, T., Norris, D.G., and de Lange, F.P.
(2016). Selective Activation of the Deep Layers of the Human Primary Visual Cortex by Top-Down Feedback. Curr. Biol. 26, 371–376.
82. Muckli, L., De Martino, F., Vizioli, L., Petro, L.S., Smith, F.W., Ugurbil, K.,
Goebel, R., and Yacoub, E. (2015). Contextual Feedback to Superficial Layers of V1. Curr. Biol. 25, 2690–2695.
83. Self, M.W., van Kerkoerle, T., Supe`r, H., and Roelfsema, P.R. (2013).
Distinct Roles of the Cortical Layers of Area V1 in Figure-Ground Segregation. Curr. Biol. 23, 2121–2129.
84. Van Kerkoerle, T., Self, M.W., and Roelfsema, P.R. (2017). Layer-specificity in the effects of attention and working memory on activity in primary visual cortex. Nat. Commun. 8, 13804.
85. Gao, Z., Davis, C., Thomas, A.M., Economo, M.N., Abrego, A.M., Svoboda, K., De Zeeuw, C.I., and Li, N. (2018). A cortico-cerebellar loop for motor planning. Nature 563, 113–116.
86. Guo, Z.V., Inagaki, H.K., Daie, K., Druckmann, S., Gerfen, C.R., and Svoboda, K. (2017). Maintenance of persistent activity in a frontal thalamocortical loop. Nature 545, 181–186.
87. Markov, N.T., Ercsey-Ravasz, M., Van Essen, D.C., Knoblauch, K., Toroczkai, Z., and Kennedy, H. (2013). Cortical High-Density Counterstream Architectures. Science 342, 1238406.
88. Tononi, G., and Edelman, G.M. (1998). Consciousness and Complexity. Science 282, 1846–1851.
89. Tononi, G. (2001). Information measures for conscious experience. Arch. Ital. Biol. 139, 367–371.
90. Tononi, G., and Sporns, O. (2003). Measuring information integration. BMC Neurosci. 4, 31.
91. Mediano, P.A.M., Rosas, F.E., Bor, D., Seth, A.K., and Barrett, A.B.(2022). The strength of weak integrated information theory. Trends Cogn. Sci. 26, 646–655.
92. Massimini, M., Boly, M., Casali, A., Rosanova, M., and Tononi, G. (2009). A perturbational approach for evaluating the brain’s capacity for consciousness. In Progress in Brain Research Coma Science: Clinical and Ethical Implications, S. Laureys, N.D. Schiff, and A.M. Owen, eds. (Elsevier), pp. 201–214.
93. Tononi, G. (2008). Consciousness as integrated information: a provisional manifesto. Biol. Bull. 215, 216–242.
94. Oizumi, M., Albantakis, L., and Tononi, G. (2014). From the Phenomenology to the Mechanisms of Consciousness: Integrated Information Theory
3.0. PLoS Comput. Biol. 10, e1003588.
95. Albantakis, L., Barbosa, L., Findlay, G., Grasso, M., Haun, A.M., Marshall,
W., Mayner, W.G.P., Zaeemzadeh, A., Boly, M., Juel, B.E., et al. (2023).
Integrated information theory (IIT) 4.0: Formulating the properties of
phenomenal existence in physical terms. PLoS Comput Biol. 19,
96. Grasso, M., Haun, A.M., and Tononi, G. (2021). Of maps and grids. Neurosci. Conscious. 2021, niab022.
97. Haun, A., and Tononi, G. (2019). Why Does Space Feel the Way it Does?
Towards a Principled Account of Spatial Experience. Entropy 21, 1160.
98. Lamme, V.A.F. (1995). The neurophysiology of figure-ground segregation
in primary visual cortex. J. Neurosci. 15, 1605–1615.
99. Lamme, V.A.F., and Roelfsema, P.R. (2000). The distinct modes of vision
offered by feedforward and recurrent processing. Trends Neurosci. 23,
100. Zipser, K., Lamme, V.A.F., and Schiller, P.H. (1996). Contextual modulation in primary visual cortex. J. Neurosci. 16, 7376–7389.
101. Lamme, V.A.F. (2015). The Crack of Dawn: Perceptual Functions and
Neural Mechanisms that Mark the Transition from Unconscious Processing to Conscious Vision (Open Mind).
102. Lamme, V.A.F. (2018). Challenges for theories of consciousness: seeing
or knowing, the missing ingredient and how to deal with panpsychism.
Philos. Trans. R. Soc. Lond. B Biol. Sci. 373, 20170344.
103. Graziano, M.S.A., and Webb, T.W. (2015). The attention schema theory: a
mechanistic account of subjective awareness. Front. Psychol. 6, 500.
104. Koivisto, M., and Revonsuo, A. (2010). Event-related brain potential correlates of visual awareness. Neurosci. Biobehav. Rev. 34, 922–934.
105. Lamme, V.A.F. (2010). How neuroscience will change our view on consciousness. Cogn. Neurosci. 1, 204–220.
106. Dehaene, S., Changeux, J.P., Naccache, L., Sackur, J., and Sergent, C.
(2006). Conscious, preconscious, and subliminal processing: a testable
taxonomy. Trends Cogn. Sci. 10, 204–211.
107. Milner, A.D., and Goodale, M.A. (1995). The Visual Brain in Action (Oxford
108. Sperling, G. (1960). The information available in brief visual presentations.
Psychol. Monogr.: Gen. Appl. 74, 1–29.
109. Self, M.W., Kooijmans, R.N., Supe`r, H., Lamme, V.A.F., and Roelfsema,
P.R. (2012). Different glutamate receptors convey feedforward and recurrent processing in macaque V1. Proc. Natl. Acad. Sci. USA 109,
110. Hohwy, J. (2013). The Predictive Mind (OUP Oxford).
111. Friston, K. (2010). The free-energy principle: a unified brain theory? Nat.
Rev. Neurosci. 11, 127–138.
112. Pennartz, C.M.A., Oude Lohuis, M.N., and Olcese, U. (2023). How ‘visual’
is the visual cortex? The interactions between the visual cortex and other
sensory, motivational and motor systems as enabling factors for visual
perception. Philos. Trans. R. Soc. Lond. B Biol. Sci. 378, 20220336.
113. Allen, W.E., Kauvar, I.V., Chen, M.Z., Richman, E.B., Yang, S.J., Chan, K.,
Gradinaru, V., Deverman, B.E., Luo, L., and Deisseroth, K. (2017). Global
Representations of Goal-Directed Behavior in Distinct Cell Types of
Mouse Neocortex. Neuron 94, 891–907.e6.
114. Oude Lohuis, M.N., Pie, J.L., Marchesi, P., Montijn, J.S., de Kock, C.P.J.,
Pennartz, C.M.A., and Olcese, U. (2022). Multisensory task demands
temporally extend the causal requirement for visual cortex in perception.
115. Oude Lohuis, M.N., Marchesi, P., Olcese, U., and Pennartz, C.M.A.
(2024). Triple dissociation of visual, auditory and motor processing in
mouse primary visual cortex. Nat. Neurosci. https://doi.org/10.1038/
116. Brucklacher, M., Bohte´ , S.M., Mejias, J.F., and Pennartz, C.M.A. (2023).
Local minimization of prediction errors drives learning of invariant object
representations in a generative network model of visual perception.
Front. Comput. Neurosci. 17, 1207361.
117. Dora, S., Bohte, S.M., and Pennartz, C.M.A. (2021). Deep Gated Hebbian
Predictive Coding Accounts for Emergence of Complex Neural
Response Properties Along the Visual Cortical Hierarchy. Front. Comput.
118. Keller, G.B., and Mrsic-Flogel, T.D. (2018). Predictive Processing: A Canonical Cortical Computation. Neuron 100, 424–435.
119. Montijn, J.S., Olcese, U., and Pennartz, C.M.A. (2016). Visual Stimulus
Detection Correlates with the Consistency of Temporal Sequences within
Stereotyped Events of V1 Neuronal Population Activity. J. Neurosci. 36,
120. Larkum, M.E. (2013). A cellular mechanism for cortical associations: an
organizing principle for the cerebral cortex. Trends Neurosci. 36,
121. Larkum, M.E., Zhu, J.J., and Sakmann, B. (1999). A new cellular mechanism for coupling inputs arriving at different cortical layers. Nature 398,
122. Larkum, M.E., Nevian, T., Sandler, M., Polsky, A., and Schiller, J. (2009).
Synaptic Integration in Tuft Dendrites of Layer 5 Pyramidal Neurons: A
New Unifying Principle. Science 325, 756–760.
123. Takahashi, N., Oertner, T.G., Hegemann, P., and Larkum, M.E. (2016).
Active cortical dendrites modulate perception. Science 354, 1587–1590.
124. Takahashi, N., Ebner, C., Sigl-Glo¨ ckner, J., Moberg, S., Nierwetberg, S.,
and Larkum, M.E. (2020). Active dendritic currents gate descending
cortical outputs in perception. Nat. Neurosci. 23, 1277–1285.
125. Suzuki, M., and Larkum, M.E. (2020). General Anesthesia Decouples
Cortical Pyramidal Neurons. Cell 180, 666–676.e13.
126. Palmer, L.M., Schulz, J.M., Murphy, S.C., Ledergerber, D., Murayama,
M., and Larkum, M.E. (2012). The Cellular Basis of GABA(B)-Mediated
Interhemispheric Inhibition. Science 335, 989–993.
127. Phillips, W.A. (2023). The Cooperative Neuron: Cellular Foundations of
Mental Life (Oxford University Press).
128. Doron, G., Shin, J.N., Takahashi, N., Druke, M., Bocklisch, C., Skenderi, €
S., de Mont, L., Toumazou, M., Ledderose, J., Brecht, M., et al. (2020).
Perirhinal input to neocortical layer 1 controls learning. Science 370,
129. Shin, J.N., Doron, G., and Larkum, M.E. (2021). Memories off the top of
your head. Science 374, 538–539.
130. Aru, J., Siclari, F., Phillips, W.A., and Storm, J.F. (2020). Apical drive-A
cellular mechanism of dreaming? Neurosci. Biobehav. Rev. 119,
131. Tononi, G., and Koch, C. (2015). Consciousness: here, there and everywhere? Philos. Trans. R. Soc. Lond. B Biol. Sci. 370, 20140167.
132. Evers, K., Farisco, M., and Pennartz, C.M.A. (2024). Assessing the
commensurability of theories of consciousness: On the usefulness of
common denominators in differentiating, integrating and testing hypotheses. Conscious. Cogn. 119, 103668. https://doi.org/10.1016/j.concog.
133. Aru, J., Larkum, M.E., and Shine, J.M. (2023). The feasibility of artificial
consciousness through the lens of neuroscience. Trends Neurosci. 46,
134. Searle, J.R. (1980). Minds, brains, and programs. Behav. Brain Sci. 3,
135. Pennartz, C.M.A., Dora, S., Muckli, L., and Lorteije, J.A.M. (2019). Towards a Unified View on Pathways and Functions of Neural Recurrent
Processing. Trends Neurosci. 42, 589–603.
136. J.R. Josephson and S.G. Josephson, eds. (1994). Abductive Inference:
Computation, Philosophy, Technology (Cambridge University Press).
137. Corcoran, A.W., Hohwy, J., and Friston, K.J. (2023). Accelerating scientific progress through Bayesian adversarial collaboration. Neuron 111,
138. Suzuki, M., Pennartz, C.M.A., and Aru, J. (2023). How deep is the brain?
The shallow brain hypothesis. Nat. Rev. Neurosci. 24, 778–791.
139. Harris, J.A., Mihalas, S., Hirokawa, K.E., Whitesell, J.D., Choi, H., Bernard, A., Bohn, P., Caldejon, S., Casal, L., Cho, A., et al. (2019). Hierarchical organization of cortical and thalamic connectivity. Nature 575,
140. Changeux, J.P. (2017). Climbing Brain Levels of Organisation from Genes
to Consciousness. Trends Cogn. Sci. 21, 168–181.
141. Rao, R.P.N., and Ballard, D.H. (1999). Predictive coding in the visual cortex: a functional interpretation of some extra-classical receptive-field effects. Nat. Neurosci. 2, 79–87.
142. Sacramento, J., Ponte Costa, R., Bengio, Y., and Senn, W. (2018). Dendritic cortical microcircuits approximate the backpropagation algorithm.
Preprint at arXiv. https://doi.org/10.48550/arXiv.1810.11393.
143. Senn, W., Dold, D., Kungl, A.F., Ellenberger, B., Jordan, J., Bengio, Y.,
Sacramento, J., and Petrovici, M.A. (2023). A Neuronal Least-Action Principle for Real-Time Learning in Cortical Circuits. Preprint at bioRxiv.
https://doi.org/10.1101/2023.03.25.534198.
144. Urbanczik, R., and Senn, W. (2014). Learning by the Dendritic Prediction
of Somatic Spiking. Neuron 81, 521–528.
145. Vladimirskiy, B., Urbanczik, R., and Senn, W. (2015). Hierarchical
Novelty-Familiarity Representation in the Visual System by Modular Predictive Coding. PLoS One 10, e0144636.
146. Markram, H., Muller, E., Ramaswamy, S., Reimann, M.W., Abdellah, M.,
Sanchez, C.A., Ailamaki, A., Alonso-Nanclares, L., Antille, N., Arsever, S.,
et al. (2015). Reconstruction and Simulation of Neocortical Microcircuitry.
147. Douglas, R.J., and Martin, K.A. (1991). A functional microcircuit for cat visual cortex. J. Physiol. 440, 735–769.
148. Attinger, A., Wang, B., and Keller, G.B. (2017). Visuomotor Coupling
Shapes the Functional Development of Mouse Visual Cortex. Cell 169,
149. Green, J., Bruno, C.A., Traunmuller, L., Ding, J., Hrvatin, S., Wilson, D.E., €
Khodadad, T., Samuels, J., Greenberg, M.E., and Harvey, C.D. (2023). A
cell-type-specific error-correction signal in the posterior parietal cortex.
150. Fahrenfort, J.J., Scholte, H.S., and Lamme, V.A.F. (2007). Masking Disrupts Reentrant Processing in Human Visual Cortex. J. Cogn. Neurosci.
151. Fahrenfort, J.J., and Lamme, V.A.F. (2012). A true science of consciousness explains phenomenology: comment on Cohen and Dennett. Trends
Cogn. Sci. 16. 138–9; author reply 139.
152. Sikkens, T., Bosman, C.A., and Olcese, U. (2019). The Role of Top-Down
Modulation in Shaping Sensory Processing Across Brain States: Implications for Consciousness. Front. Syst. Neurosci. 13, 31.
153. Dehaene, S., Artiges, E., Naccache, L., Martelli, C., Viard, A., Schurhoff, €
F., Recasens, C., Martinot, M.L.P., Leboyer, M., and Martinot, J.L. (2003).
Conscious and subliminal conflicts in normal subjects and patients with
schizophrenia: The role of the anterior cingulate. Proc. Natl. Acad. Sci.
154. Zylberberg, A., Dehaene, S., Mindlin, G.B., and Sigman, M. (2009).
Neurophysiological bases of exponential sensory decay and top-down
memory retrieval: a model. Front. Comput Neurosci. 3, 4.
155. Doerig, A., Schurger, A., and Herzog, M.H. (2021). Hard criteria for empirical theories of consciousness. Cogn. Neurosci. 12, 41–62.
156. Knierim, J.J., and Van Essen, D.C. (1992). Visual cortex: cartography,
connectivity, and concurrent processing. Curr. Opin. Neurobiol. 2,
157. Laureys, S. (2005). The neural correlate of (un)awareness: lessons from
the vegetative state. Trends Cogn. Sci. 9, 556–559.
158. Hill, S., and Tononi, G. (2005). Modeling Sleep and Wakefulness in the
Thalamocortical System. J. Neurophysiol. 93, 1671–1698.
159. Laureys, S., Owen, A.M., and Schiff, N.D. (2004). Brain function in coma,
vegetative state, and related disorders. Lancet Neurol. 3, 537–546.
160. Bayne, T., Hohwy, J., and Owen, A.M. (2016). Are There Levels of Consciousness? Trends Cogn. Sci. 20, 405–413.
161. Steriade, M. (2006). Grouping of brain rhythms in corticothalamic systems. Neuroscience 137, 1087–1106.
162. Heeger, D.J. (2017). Theory of cortical function. Proc. Natl. Acad. Sci.
163. Merker, B. (2007). Consciousness without a cerebral cortex: A challenge
for neuroscience and medicine. Behav. Brain Sci. 30, 63–81.
164. Penfield, W., and Jasper, H. (1954). Epilepsy and the Functional Anatomy
of the Human Brain (Little, Brown & Co.).
165. Solms, M., and Friston, K. (2018). How and why consciousness arises:
Some considerations from physics and physiology. J. Conscious. Stud.
166. Kanai, R., Komura, Y., Shipp, S., and Friston, K. (2015). Cerebral hierarchies: predictive processing, precision and the pulvinar. Philos. Trans. R.
Soc. Lond. B Biol. Sci. 370, 20140169.
167. Pennartz, C.M.A. (2009). Identification and integration of sensory modalities: Neural basis and relation to consciousness. Conscious. Cogn. 18,
168. Luczak, A., Bartho´ , P., and Harris, K.D. (2009). Spontaneous Events
Outline the Realm of Possible Sensory Responses in Neocortical Populations. Neuron 62, 413–425.
169. Montijn, J.S., Goltstein, P.M., and Pennartz, C.M. (2015). Mouse V1 population correlates of visual detection rely on heterogeneity within
neuronal response patterns. eLife 4, e10163.
170. Demertzi, A., Tagliazucchi, E., Dehaene, S., Deco, G., Barttfeld, P., Raimondo, F., Martial, C., Ferna´ ndez-Espejo, D., Rohaut, B., Voss, H.U.,
et al. (2019). Human consciousness is supported by dynamic complex
patterns of brain signal coordination. Sci. Adv. 5, eaat7603.
171. Farisco, M., and Changeux, J.P. (2023). About the compatibility between
the perturbational complexity index and the global neuronal workspace
theory of consciousness. Neurosci. Conscious. 2023, niad016.
172. Cattani, A., Galluzzi, A., Fecchio, M., Pigorini, A., Mattia, M., and Massimini, M. (2023). Adaptation Shapes Local Cortical Reactivity: From Bifurcation Diagram and Simulations to Human Physiological and Pathological Responses. eNeuro 10, 435–22.2023.
173. Chang, J.Y., Fecchio, M., Pigorini, A., Massimini, M., Tononi, G., and Van
Veen, B.D. (2019). Assessing recurrent interactions in cortical networks:
Modeling EEG response to transcranial magnetic stimulation.
J. Neurosci. Methods 312, 93–104.
174. Pigorini, A., Sarasso, S., Proserpio, P., Szymanski, C., Arnulfo, G., Casarotto, S., Fecchio, M., Rosanova, M., Mariotti, M., Lo Russo, G., et al.
(2015). Bistability breaks-off deterministic responses to intracortical
stimulation during non-REM sleep. NeuroImage 112, 105–113.
175. Frith, C. (2005). The neural basis of hallucinations and delusions. C. R.
176. Seth, A.K. (2021). Being You: A New Science of Consciousnes. https://
books.google.nl/books/about/Being_You.html?id=7d9UygEACAAJ
177. Casarotto, S., Comanducci, A., Rosanova, M., Sarasso, S., Fecchio, M., Napolitani, M., Pigorini, A., G Casali, A., Trimarchi, P.D., Boly, M., et al. (2016). Stratification of unresponsive patients by an independently validated index of brain complexity. Ann. Neurol. 80, 718–729.
178. Comanducci, A., Casarotto, S., Rosanova, M., Derchi, C.C., Vigano` , A., Pirastru, A., Blasi, V., Cazzoli, M., Navarro, J., Edlow, B.L., et al. (2023). Unconsciousness or unresponsiveness in akinetic mutism? Insights from a multimodal longitudinal exploration. Eur. J. Neurosci. https://doi.org/ 10.1111/ejn.15994.
179. Arena, A., Juel, B.E., Comolatti, R., Thon, S., and Storm, J.F. (2022). Capacity for consciousness under ketamine anaesthesia is selectively associated with activity in posteromedial cortex in rats. Neurosci. Conscious. 2022, niac004.
180. Cavelli, M.L., Mao, R., Findlay, G., Driessen, K., Bugnon, T., Tononi, G., and Cirelli, C. (2023). Sleep/wake changes in perturbational complexity in rats and mice. iScience 26, 106186.
181. Ho¨ nigsperger, C., Storm, J.F., and Arena, A. (2023). Laminar evoked responses in mouse somatosensory cortex suggest a special role for deep layers in cortical complexity. Eur. J. Neurosci. https://doi.org/10.1111/ ejn.16108.
182. D’Andola, M., Rebollo, B., Casali, A.G., Weinert, J.F., Pigorini, A., Villa, R., Massimini, M., and Sanchez-Vives, M.V. (2018). Bistability, Causality, and Complexity in Cortical Networks: An In Vitro Perturbational Study. Cereb. Cortex 28, 2233–2242.
183. Gidon, A., Aru, J., and Larkum, M.E. (2022). Does brain activity cause consciousness? A thought experiment. PLoS Biol. 20, e3001651.
184. Searle, J.R. (2013). Can Information Theory Explain Consciousness? N. Y. Rev. Books 60.
185. Tononi, G., and Koch, C. (2013). Can a Photodiode Be Conscious? N. Y. Rev. Books 60.
186. Tononi, G., Albantakis, L., Boly, M., Cirelli, C., and Koch, C. (2023). Only What Exists Can Cause: an Intrinsic View of Free Will. arXiv. https://doi. org/10.48550/arXiv.2206.02069.
187. Fleming, S., Frith, C., Goodale, M., Lau, H., LeDoux, J.E., Lee, A.L.F., Michel, M., Owen, A., Peters, M.A.K., and Slagter, H.A. (2023). The Integrated Information Theory of Consciousness as Pseudoscience. Preprint at PsyArXiv. https://doi.org/10.31234/osf.io/zsr78. 188. Whyte, C.J., and Smith, R. (2021). The predictive global neuronal workspace: A formal active inference model of visual consciousness. Prog. Neurobiol. 199, 101918.
189. Hohwy, J., Tononi, G., Seth, A.K., and Tsuchiya, N. (2015). Can the free energy principle be used to generate a theory of consciousness? In Conference Abstract: XII International Conference on Cognitive Neuroscience (ICON-XII). https://doi.org/10.3389/conf.fnhum.2015.217.00420.
190. Storm, J.F. (2020). Why Does the Brain-Mind (Consciousness) Problem Seem So Hard? J. Conscious. Stud. 27, 174–189.
191. Farisco, M., Evers, K., Annen, J., Blandin, V., Camassa, A., Cecconi, B., Charland, V., Deco, G., Gosseries, O., Laureys, S., et al. (2023). Advancing the Science of Consciousness: from Ethics to Clinical Care. Preprint at PsyArXiv. https://doi.org/10.31234/osf.io/sutrc.
192. Massimini, M., Ferrarelli, F., Huber, R., Esser, S.K., Singh, H., and Tononi, G. (2005). Breakdown of Cortical Effective Connectivity During Sleep. Science 309, 2228–2232.
193. Olcese, U., Oude Lohuis, M.N., and Pennartz, C.M.A. (2018). Sensory Processing Across Conscious and Nonconscious Brain States: From Single Neurons to Distributed Networks for Inferential Representation. Front. Syst. Neurosci. 12, 49.
194. Suzuki, M., and Larkum, M.E. (2017). Dendritic calcium spikes are clearly detectable at the cortical surface. Nat. Commun. 8, 276.
195. Goltstein, P.M., Montijn, J.S., and Pennartz, C.M.A. (2015). Effects of Isoflurane Anesthesia on Ensemble Patterns of Ca2+ Activity in Mouse V1: Reduced Direction Selectivity Independent of Increased Correlations in Cellular Activity. PLoS One 10, e0118277.
196. Albantakis, L., Hintze, A., Koch, C., Adami, C., and Tononi, G. (2014). Evolution of Integrated Causal Structures in Animats Exposed to Environments of Increasing Complexity. PLoS Comput Biol. 10, e1003966.
197. Deperrois, N., Petrovici, M.A., Senn, W., and Jordan, J. (2022). Learning cortical representations through perturbed and adversarial dreaming. eLife 11, e76384.
198. O’Connor, D.H., Hires, S.A., Guo, Z.V., Li, N., Yu, J., Sun, Q.Q., Huber, D., and Svoboda, K. (2013). Neural coding during active somatosensation revealed using illusory touch. Nat. Neurosci. 16, 958–965. 199. Storm, J.F., Boly, M., Casali, A.G., Massimini, M., Olcese, U., Pennartz,
C.M.A., and Wilke, M. (2017). Consciousness Regained: Disentangling Mechanisms, Brain Systems, and Behavioral Responses. J. Neurosci.37, 10882–10893.
什么是意识?意识能否度量?机器能否产生意识?对于意识问题,人们可能即将迎来一个大的突破,各种有关意识的理论正如雨后春笋般展现出勃勃生机。其中神经科学家 Giulio Tononi 的整合信息论(IIT)被认为是最有前景的意识理论之一。如果说意识是大脑神经活动的一种涌现结果,那么刻画涌现便成为理解意识过程中一个重要环节。因果涌现理论目前发展出两个派别,除了 Erik Hoel 的有效信息因果涌现框架,还有一个是 Rosas 的信息分解(PID)框架,此后 Rosas 基于此进一步提出融合整合信息论的信息分解框架 ΦID,尝试构建新的意识理论。
一边是信息整合(IIT),一边是信息分解(PID),看似分裂,实际上都是对香农经典信息论的进一步发展。因果涌现读书会第四季「新信息论:从分解到整合」由北京师范大学系统科学学院教授、集智俱乐部创始人张江老师领衔发起,旨在梳理信息论领域的发展脉络,从香农的经典信息论开始,重点关注整合信息论和信息分解这两个前沿话题,及其在交叉领域的应用。希望通过对这些“新信息论”度量指标的深入探讨,帮助我们理解什么是意识,什么是涌现,并找到不同学科,不同问题背后的统一性原理。