Nature Methods:综述回顾哺乳动物细胞中的定向进化技术:核心突变方法盘点解析
导语
定向进化(directed evolution),是指在实验室的特定条件下对自然进化的模拟和改进。
2018 年,因为其在 “酶的定向进化” 领域上所做出的贡献,弗朗西斯・阿诺德(Frances H. Arnold)教授摘得了当年的诺贝尔化学奖。正如获奖理由中所说那样:这项技术(定向进化)帮助了科学家们 “掌控了进化的力量 ”。

王韵晴 | 作者
生辉SynBio | 来源

论文标题:
Directed evolution in mammalian cells
论文地址:
https://www.nature.com/articles/s41592-021-01090-x

图丨 2018 年诺贝尔化学奖得主(来源:Nobel Prize)
在自然过程中,物种的繁殖会伴随有少数随机产生的变异,一旦生存环境发生改变,而变异者比非变异者更适应这种变化的话,那么它们将会 “被环境选择” 而得以存活,从而实现了物种的进化。
与之类似,要在定向进化过程中复现这一过程,首先便是在目标蛋白的基因水平上尽可能多地人为诱发突变,形成一个庞大的基因突变库,然后进行表达,得到相应的突变库。最终,只有符合研究人员需求的具有特定性质的突变体会 “脱颖而出”。
值得关注的是,定向进化技术能使基因突变更迅速,后续选择更高效,这样可以让蛋白质尽快达到预期的表现形式或者功能状态。如今,科学家们已借助 “试管里的达尔文进化论” 得到了性能优化的酶、荧光蛋白、离子通道、抗体等等。
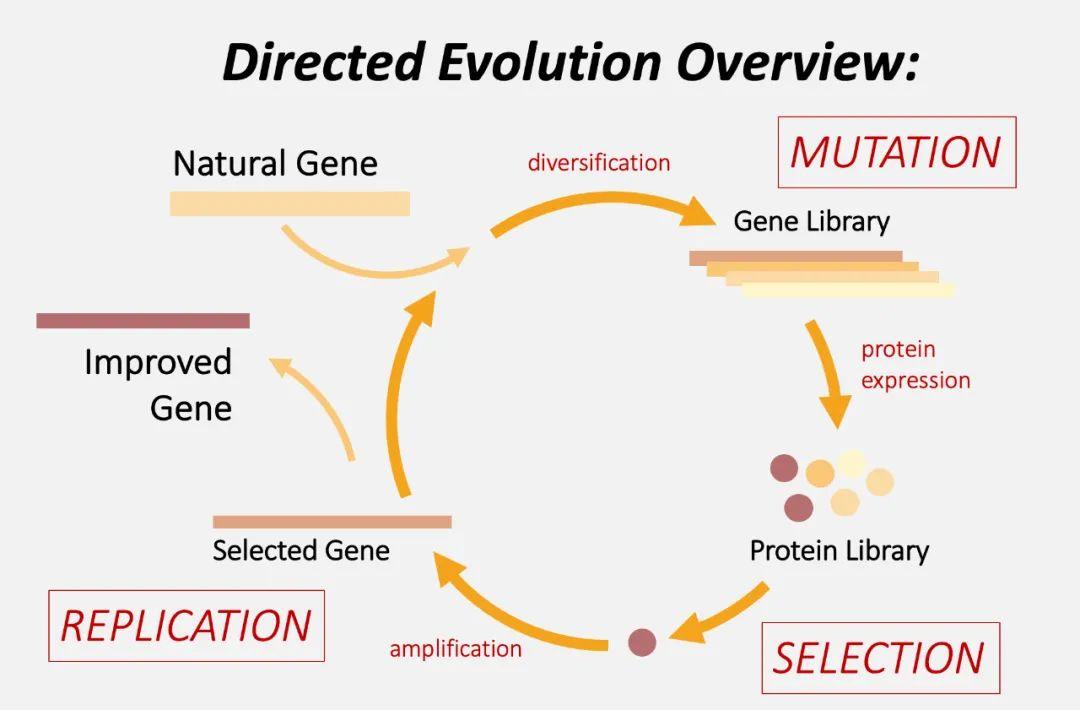
图丨定向进化流程示意图(图源:Stanford)
定向进化技术发展前期,大多致力于改造酶的特异性、稳定性、催化活性等等,服务于能源化工领域,因此在试管里或者细菌酵母里完成定向进化。然而,通过此类技术路线得到的蛋白质,在应用到生物学和医学领域时,往往无法达到预期效果。
此外,在非哺乳动物细胞环境下进化得到的蛋白质,常常会因错误的折叠修饰、分子间相互作用、错误的细胞定位等因素,使之无法在复杂的哺乳动物细胞中正常工作。
因此,开发能够应用于哺乳动物细胞中的定向进化技术,便是一大趋势。
而就在 4 月 7 日,来自 MIT 化学系的 Samuel J. Hendel 和 Matthew D. Shoulders 副教授在 Nature Methods 发表了相关综述,对在哺乳动物细胞中实现定向进化的研究进展和困难挑战进行了梳理。
图丨 适用于哺乳动物细胞的定向进化(图源:Nature Methods)
1.突变 – 表达 – 筛选或选择
1.突变 – 表达 – 筛选或选择
要完成一轮定向进化,通常需要分为三步:“突变”、“表达” 以及 “筛选或选择”。
第一步便是构建目的基因的突变库。人工引入突变的方法一般分为细胞外突变法(ex mammalia mutagenesis)和细胞内突变法(in mammalia mutagenesis)。
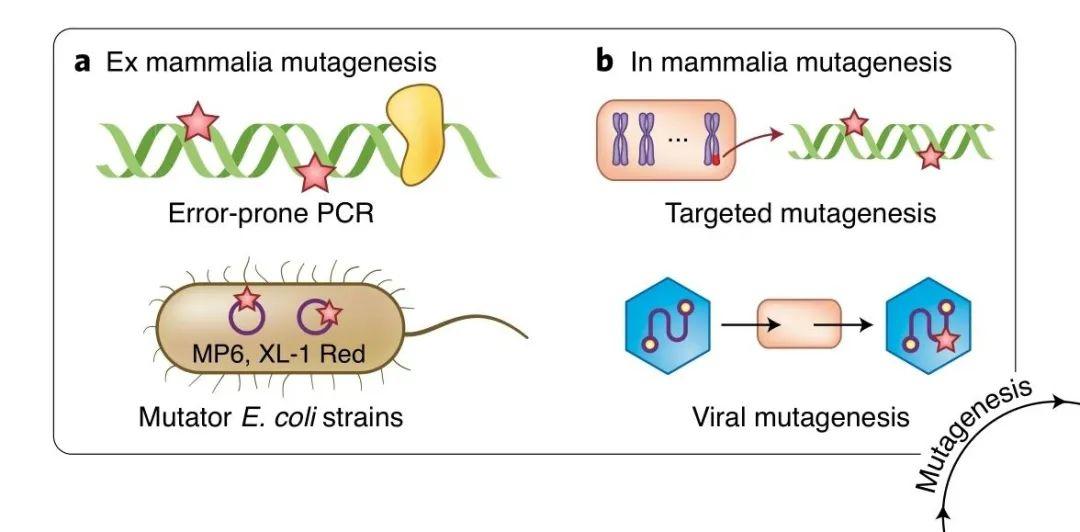
图丨 典型的突变法(图源:Nature Methods)
细胞外突变法主要包括:可以产生随机突变的的易错 PCR(error-prone PCR)和易突变菌株(mutator strains of bacteria),以及可以产生非随机突变的定点突变(site-directed mutagenesis)和合成文库(synthetic library generation)。
细胞内突变法主要包括:体细胞超突变(somatic hypermutation)、基于 CRISPR 的定点突变(CRISPR-based DNA targeting)、基于 RNA 聚合酶的持续突变( highly processive RNA polymerases)和病毒辅助的突变(viral replication)。
突变是发生在基因层面上的,而筛选或选择需要通过表型进行,因此便需要一个中间环节将两者联系起来,那便是:“表达”。通常研究人员建立基因突变文库后,需要先在细胞内瞬时地或者稳定地表达生成蛋白质,才能够进行下一步的挑选和验证。
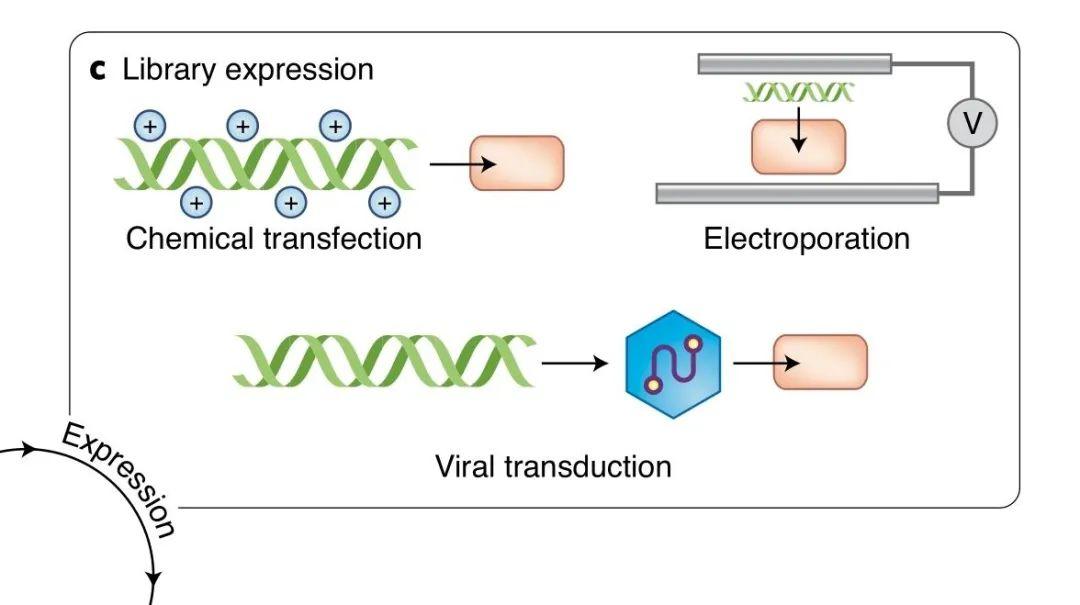
图丨 典型的表达法(图源:Nature Methods)
瞬时表达法中,包括化学转染法和电转染法,转染效率、细胞间不一致的表达水平、多突变体在一个细胞中表达和细胞系兼容性,都是干扰实验的因素。
稳定表达法中,包括逆转录病毒转导法(retroviral transduction)和定点重组法(site-specific recombination),解决了前者存在的一些干扰因素,尤其能基本保障细胞间表达水平一致。
最后一步,便是研究人员对细胞内表达的突变体进行评估,挑选出符合目标功能需求的进化突变体。
在挑选过程中,有着两种策略:一种是筛选(screening),另一种是选择(selection)。二者的区别在于,筛选通常以物理性质为判断依据(例如光学性质),而选择通常以生物表型为判断依据(例如细胞活性)。
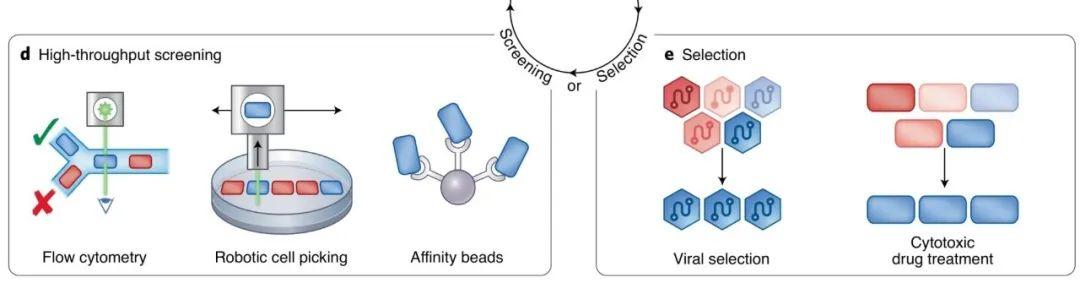
图丨 典型的筛选法和选择法(图源:Nature Methods)
在筛选法中,研究人员会给目的蛋白的功能设定一个阈值,然后通过荧光流式细胞术(fluorescence-based flow cytometry)或细胞显示法(cellular display),机械性地分离出高出阈值的、具备理想突变体的细胞。
而在选择法中,研究人员会设计合理的细胞调控回路,使得目的蛋白定向进化出的功能,能显著影响细胞存活的必要因素。随后人为施加生存环境上的选择压力,使得只有具备特定性质的 “适合环境” 的突变体才能够存活。
筛选法需要研究人员手动处理每轮定向进化之前、之中和之后的细胞,以及确定每一轮的筛选阈值,人力成本较高。相比之下,选择法对人员和设备的要求更低、更易于实现规模化和多次迭代。
2.突变方法盘点
2.突变方法盘点
自定向进化技术问世以来,已经衍生出相当多的细分技术。在各类技术之中,可控制、高效率的突变一直是定向进化的核心关键。
正如前文所述,在哺乳细胞中的突变方法通常可以分为两类:细胞外突变法和细胞内突变法。而在细胞内突变法之中,新兴的病毒辅助方法则进一步拓展了定向进化的应用范围。
细胞外突变法
细胞外突变法是最为成熟且直观的定向进化方法,通过该方法与不同种类的筛选方法的联用,研究人员已经成功进化出了许多有价值的蛋白质,比如:不稳定结构域、离子通道及致癌蛋白等等。
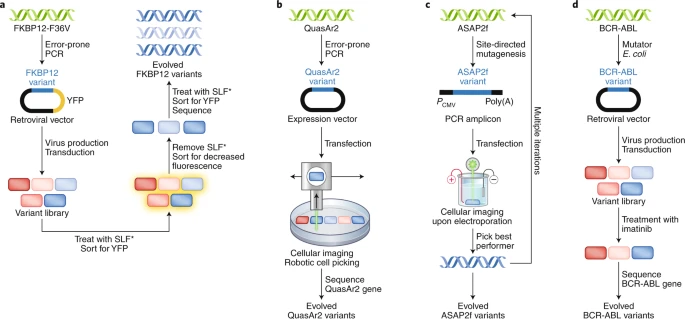
图丨 利用细胞外突变法实现定向进化的例子(图源:Nature Methods)
利用细胞外突变结合筛选法,可以进化出能被特异性小分子配体调控的不稳定结构域,那是一种控制功能性蛋白的稳定性的分子工具。而如果使用其结合选择法的话,则通常被用于阐明致癌蛋白是如何产生耐药性的。
虽然细胞外突变法成熟且直观,不过其存在着严重的缺陷:由于突变库的表达和提取需要密集的人工操作,在大多数情况下,细胞外突变法只能在相当有限的突变库上进行一轮进化。
然而,为了得到更有优势的蛋白质,或者达成更复杂的定向进化目标,就需要更大体量的突变库以及执行多轮迭代的进化。
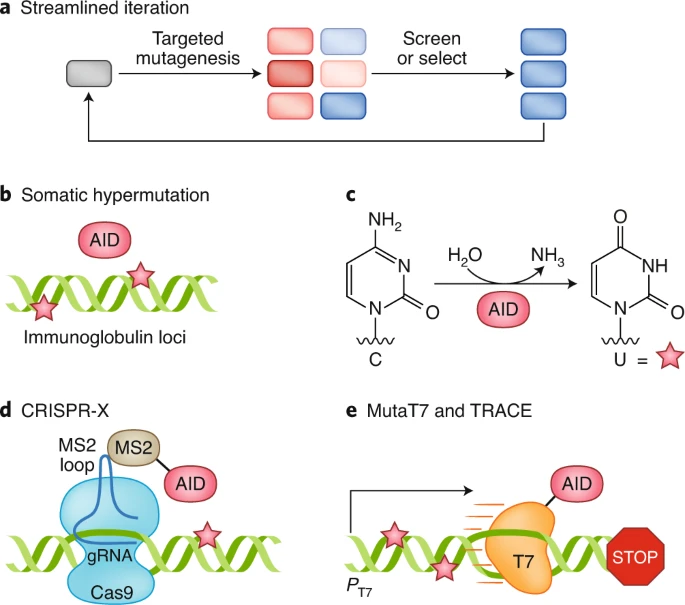
图丨 可迭代的定向进化示意图(图源:Nature Methods)
近五年里,研究人员们致力于突破这个瓶颈,开发了细胞内突变法。
细胞内突变法
相较于微生物,在哺乳动物细胞里进行细胞内突变法显然更具挑战性。哺乳动物细胞基因组规模巨大,这意味着,更容易发生一些能欺骗筛选或选择策略的脱靶突变。
为了更加快速、高效、准确地实现细胞内的连续进化,研究人员们不断对目前已有的细胞内突变法进行优化和改进。
图丨 利用细胞内突变法实现定向进化的例子(图源:Nature Methods)
目前在细胞内突变法中,常用的诱发突变的分子工具是脱氨酶。
活化诱导的胞苷脱氨酶(activation-induced cytidine deaminase,AID),能将胞嘧啶脱氨转化为尿嘧啶。由此产生的尿嘧啶,可能被错误修复,也可能在复制时错误与腺嘌呤而不是鸟嘌呤结合,因而诱发突变。
基于此,研究人员实现了对哺乳动物细胞中的抗体和荧光蛋白的定向进化。
可惜的是,AID 并非只作用于特定基因位点,而是会普遍地在高转录区域诱导基因突变。那么,我们就需要给 “激光” 装上 “定位系统”。
现有的 “定位系统” 主要是 Cas 酶和 T7 RNA 聚合酶。
Cas9 可在向导 RNA(guide RNA)的引导下靶向切割基因组位点。进行切割以后,细胞会以两种机制修复此位点,非同源末端连接(Non-homologous End Joining)或者同源定向修复(Homology directed repair),诱发突变。
基于此,研究人员得到了具有耐药性突变的蛋白,和在低 pH 环境的溶酶体中也能维持生理活性的荧光蛋白。
新兴的 CRISPR 工具诚然实现了定点引入突变的突破,但是,如果想要实现多轮迭代的进化,就需要能识别积累了多次突变的位点的 gRNA。此外,该技术仍然受到切割位点范围的限制,仅在 gRNA 互补区附近 (~10–50 bp) ,而目的基因不一定具备足够多的合适位点。
为了解决突变窗口小这一问题,研究人员引入了和 Cas9 同样具备靶向能力的,而且定位范围更广的 RNA 聚合酶。T7 RNA 聚合酶(T7 RNA polymerase)识别特异性的 T7 启动子后,并且能够覆盖超过 10000 bp 的 DNA。
研究人员结合 T7 RNA 聚合酶与胞苷脱氨酶(cytidine deaminase),开发了 TRACE (T7 polymerase-driven continuous editing) 系统,实现特定基因的哺乳动物细胞内连续进化。利用 TRACE,研究人员已筛选出在 A375 细胞中对小分子抑制剂具有抗药性的 MEK 变种。
从理论上讲,现在可以很容易地将 RNA 引导的核酸内切酶或 T7 聚合酶直接在哺乳动物细胞中瞄准的任何基因变异。
尽管如此,上述方法仍受到诸多因素的制约。首先,哺乳动物细胞生长速度缓慢。其次,细胞能自发通过其他旁路途径获得对选择压力的抵抗力。再者,上述方法只关注了优化建立突变库的策略,而仍然依赖于传统的选择或筛选方法,可选择的具备可观察的表型或与细胞活性相关的生物分子的范围较为狭窄。
而最近开发的病毒辅助定向进化平台,克服了上述基于细胞的方法的许多局限性,使得可以通过定向进化来实现功能优化的生物分子的范围大大扩大。
病毒辅助法
病毒辅助定向进化的概念很简单:将感兴趣的基因引入病毒的基因组中,然后将病毒在细胞中繁殖的能力与定向进化目标蛋白的功能联系在一起,病毒以低保真度复制。这就意味着,那些产生符合需求的突变的病毒,大量繁殖,成为优势群体。
在开发适用于哺乳动物细胞的病毒辅助定向进化技术时,研究人员需要考虑的一般原则包括:病毒的复制速度不能超过目的基因的表达速度,尽量选择宿主广泛的病毒,以及病毒生产包装的实用性和在实验室操作的安全性。
两个里程碑式的工作,极大促进了病毒辅助定向进化的发展。
一个是 Matthew D. Shoulders 实验室在 2018 年开发的适用于哺乳动物的噬菌体辅助连续进化系统(mammalian Phage-Assisted Continuous Evolution,mPACE),另一个是 Bryan L. Ruth 实验室在 2019 年开发的基于病毒的精准序列进化系统(viral evolution of genetically actuating sequences,VEGAS)。
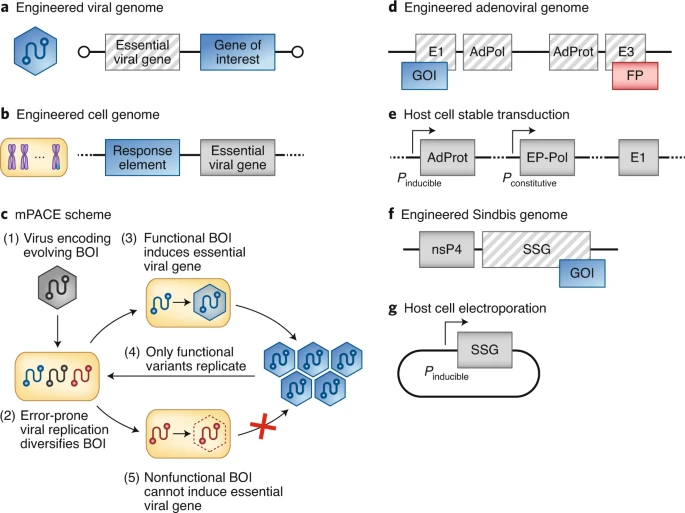
图丨病毒辅助定向进化的设计流程图(图源:Nature Methods)
mPACE 和 VEGAS 的基本原理相近:首先,删除病毒基因组里的对于病毒生存必须的基因,同时引入感兴趣的目的基因;接着,改造宿主哺乳动物细胞的基因组,把想要进得到的目的基因的功能与原本缺失的病毒基因的转录、翻译或激活,偶联在一起。
这样一来,只有进化出我们感兴趣功能的病毒突变株,才能被分泌到细胞外,感染新的细胞并进行下一轮复制增殖,数量增多,从而在进化池中保留下来,实现了生物体 “自发” 的连续定向进化。
相比于其它技术,病毒辅助定向进化技术可谓是取得了重大突破。其显著优势体现在速度快,人力劳动成本低 ,避免了哺乳动物基因组中脱靶突变的积累,以病毒丰度为选择依据,为更多目的蛋白的进化提供可能。
它能提供最大的突变库和最短的实验时间尺度,原则上允许最广泛的生物分子进化,极大地帮助研究人员获得更丰富的更理想的蛋白质。
3.定向进化的潜力
3.定向进化的潜力
总结来说,目前在哺乳动物之中成功进化的蛋白质大致分为三类:参与细胞保守的生理过程、多细胞生物特有的生理过程、或者哺乳动物特有的生理过程,很显然,基于哺乳动物细胞的定向进化技术对于研究和改造这三类蛋白是必不可少的。
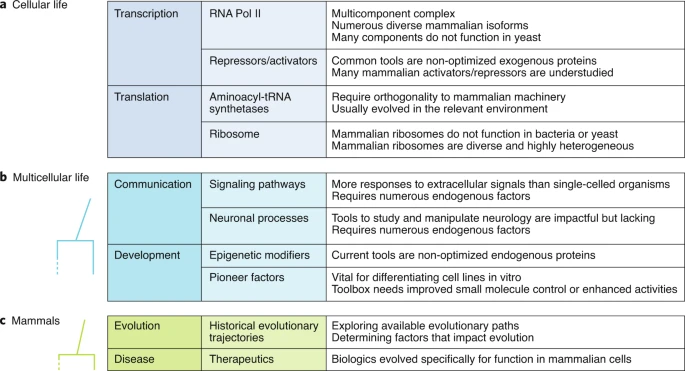
图丨通过定向进化得到的适用于哺乳动物细胞的蛋白进展总结(图源:Nature Methods)
通过在实验室模拟并加速自然的进化过程,我们不仅可以针对某目标蛋白进行改造,还可以优化细胞代谢网络、获得期望表型的细胞,为合成生物学开发更丰富更精确的功能元件、设计组装系统提供了强有力的工具。
定向进化技术,在工业生产、临床医学和基础科研方面都蕴藏着巨大的潜力。我们可以通过定向进化,获得性能优异的蛋白质,应用于能源、化工、医疗等领域。通过对定向进化结果的分析,我们也更深刻地探索蛋白质从序列到结构到功能间的微妙关系。
“适者生存没有一定之规,只有保持变化才是生命的长青之道。” 随着定向进化技术变得更加高效和普适,研究人员在运用进化的力量上,愈发得心应手。
复杂科学最新论文
集智斑图顶刊论文速递栏目上线以来,持续收录来自Nature、Science等顶刊的最新论文,追踪复杂系统、网络科学、计算社会科学等领域的前沿进展。现在正式推出订阅功能,每周通过微信服务号「集智斑图」推送论文信息。扫描下方二维码即可一键订阅:

推荐阅读
点击“阅读原文”,追踪复杂科学顶刊论文
微信扫一扫,分享到朋友圈











