周昌松 | 作者
懿章 | 整理
邓一雪 | 编辑
1.6.2两种模型
2.1基于电导的神经网络模型
第三部分:少即是多,神经回路动力系统的成本效益
第四部分:学习、记忆等大脑自组织活动
第五部分:恢复清醒过程中皮层波的传播
第六部分:大尺度脑结构的功能特征处理
第七部分:基于机器学习的神经计算模型
大脑作为一个复杂系统,不仅处理复杂的外界信息,内部也有自组织的复杂的脑活动。而我们对内部的这种自组织活动的机理及其功能意义知之甚少,周昌松老师对相关的研究进行概览性的介绍,为想入门神经动力学领域的朋友提供一个入门路径。
作为一个多尺度的复杂动力学网络系统和功能系统,神经系统具有多方面的显著特征,特别是在成本效益均衡和建模方面。本文会从大脑活动的生物物理学特性到动力学模型构建等角度,简要地介绍复杂神经动力学研究基础。
大脑具有显著的多尺度特征。在神经元尺度,具有轴突、树突等结构形成的局部回路;在毫米尺度具有层和柱结构。局部回路在大脑皮层表面形成灰质,内部的白质是不同脑区轴突形成的纤维,像高速公路一样把不同的脑区连接起来,从而赋予大脑高级功能。单个神经元的作用是产生1ms左右的短时动作电位,以在神经网络中进行传输和计算。
一个成人的大脑大约具有860亿个神经元,每个神经元有成千上万量级的突触连接来与其它神经元沟通,这也造就了复杂的大脑活动。大脑的重量占体重的2%,消耗能量却占身体总耗能的20%,也侧面说明了大脑活动的复杂性和功能的重要性。但是大脑处理如此复杂的活动,消耗的能量却只相当于一个20w的灯泡,说明大脑活动是极其高效的。
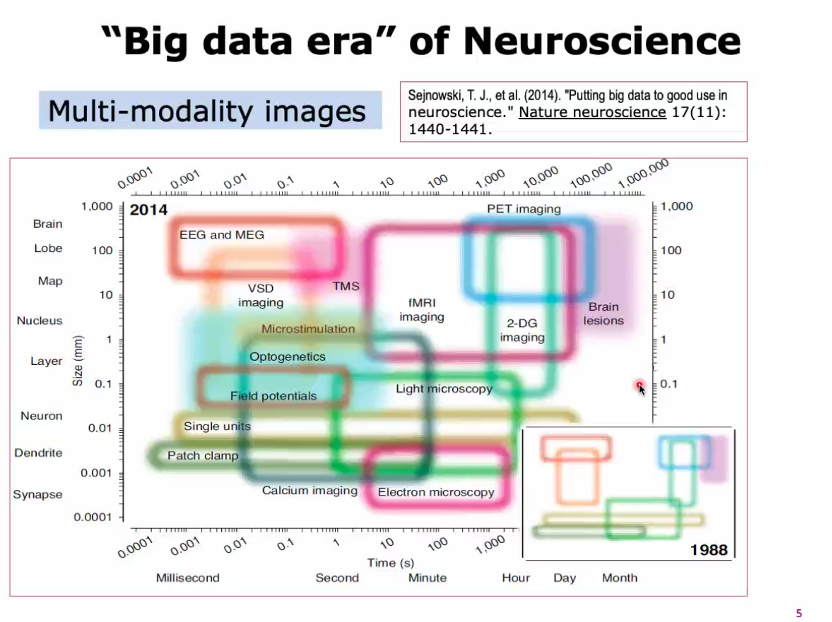
大脑作为一个时间、空间多尺度的复杂系统,可供神经科学研究的数据量极为丰富。神经科学家们可以基于研究兴趣选择不用的视角与方法。
脑电图(Electroencephalography, EEG):EEG 测量由大脑神经元内的离子电流引起的电压波动,最常用于诊断癫痫。通常侧重于测量事件相关电位或脑电波的类型,可提供毫秒级时间分辨率。
功能磁共振成像(Functional magnetic resonance imaging, fMRI):通过检测血流的变化(血氧水平依赖信号,blood-oxygen-level dependent, BOLD)来测量大脑活动。这种技术测量原理是当大脑的某个区域兴奋时,流向该区域的血流量也会增加,主要反映了神经元的输入和神经元内的信息处理,而较少反映神经元的输出放电。
从时间空间尺度来看,EEG可以实现无创全脑测量,空间精度差,时间精度高;fMRI可以实现毫米级的大脑活动测量,但是通过测量血氧含量间接反应大脑活动。对于动物如小鼠,还可以通过转基因技术,测量皮层示光变化以测量电压变化,时间、空间精度都很高[2]。
世界范围内正在陆续实施脑计划,如欧洲的HCP (The Human Connectome Project) 或是近年开展的“中国脑计划”,均希望更好地理解大脑的功能与障碍,并促进类脑、脑启发的计算与硬件计算机的发展。
对于大脑活动而言,即使在静息状态下它也有很多自发的活动,这表明复杂动力学是行为的基础。而有研究显示,大脑的自发复杂动力学的耗能占大脑整体耗能的95%,这是为什么呢?可以推测,大脑复杂动力学具有重要的功能意义。但在认知科学等的研究中,从EEG平均得到ERP(事件相关电位), 自发复杂动力学通常会被当做随机过程忽略了。
大脑的行为不能轻易地从它的属性中推断出来,表现出的复杂性具有很重要的功能研究价值。因此使用一些数学物理上的手段去描述大脑的活动,可以帮助我们探寻大脑深层次的机制。
我们以一种有效的方法举例。多尺度熵(Multiscale entropy, MSE)是一种衡量时间序列复杂性有效的方法,可直接提取原始信号包含的模式信息。该方法通过测量神经动力学在不同时间尺度的波动复现概率,刻画复杂性[3]。这种方法在生物系统、地球科学和机械振动方面都显示出良好的效果。
目前已有许多研究利用动力学模型刻画不同的脑疾病,如:
1.注意缺陷多动障碍复杂性的刻画[4,5]。
2.抑郁[6]
3.精神分裂[7]
非线性加上高自由度就成就了复杂性。神经元具有超强的非线性,而数量惊人。大脑连接组作为网络的网络,可以使用复杂网络的一些表征,例如小世界、无标度(scale-free,即网络的度分布遵循幂律)、模块化(modular,即高度模块化的网络在模块内的节点之间具有密集的连接,但在不同模块中的节点之间具有稀疏的连接)、层次性(hierarchy,即如同“树”的生成,节点与节点之间的连接体现了一种分层结构)等来刻画。
大脑网络连接突出的特点是不均匀,一些神经元之间或脑区之间的连接更紧密,与其它神经元或脑区之间的连接更松散,形成模块结构, 连接随距离衰减,这是一种功能的安排,为了实现高效的活动处理, 也可以节省能量[9,10]。
由于神经元具有强非线性,因此在单个神经元层次,神经元的活动是极其混沌和随机的,但将所有神经元整合起来的系统具有自发涌现的活动[11]。
在神经元尺度,神经动力学具有低发放率、不规则的特征[12,13];但是为什么会出现无规则的发放?从兴奋-抑制(E-I)平衡的角度理解,即兴奋神经元和抑制神经元的互相作用达到平衡,理论上导致随机非同步发放[14-16]。
而随着研究对兴奋-抑制神经元更细致的测量,发现他们并非完全的不同步[17]。在更大空间尺度上,神经动力学又会具有临界雪崩、多时间尺度的网络震荡和空间上皮层波的传播等现象。
神经元集群尺度
神经元集群发放的概率并不低,但成对神经元的关联性几乎为0。但如果将神经元打乱,集群发放的概率会显著下降。有研究显示:如果使用Ising模型,原则上使用随机网络的神经元集群(几十个)用微弱的成对关联性也可以产生较高概率的集群发放[18]。对于毫米量级的神经元集群,也会出现大范围的同时发放,该现象被称做雪崩。这种活动出现的概率服从无标度的幂律分布[19,20]。
当然,也有研究运用局域场方法测量电势,观察雪崩是否扩展[21-23]。
全皮层尺度
如下研究从全脑皮层的尺度上研究了不同状态下小鼠或人类大脑的雪崩现象:小鼠静息态(麻醉-清醒)[24,25] ,小鼠任务态(看电影)[26],人脑静息态fMRI, 从小的尺度到全脑都可以发现服从幂律分布的复杂性活动[27,28]。
在物理学上幂律分布与临界性、相变有关,类似于自旋铁磁体的临界性,即在温度升高到一定值后,磁性消失,波动变化服从幂律分布。在大脑活动研究中可以理解为自组织临界性,无需细调参数,因此具有一定争议[29]。
有外界刺激时
当有外界刺激时,大脑会进行响应。临界性会发生偏移,对外界的刺激会相较变得不敏感[30,32]。这种系统可以对外界刺激随时做出响应,但是需要消耗大量的能量的大脑临界态可以可以临界态解释大脑“暗能量”,即需要大量的能量构建大脑复杂自发动态系统。
神经振荡是中枢神经系统中神经活动的一种规律。神经组织可以通过多种方式产生振荡活动,由单个神经元内的机制或神经元之间的相互作用驱动。在单个神经元中,振荡可以表现为膜电位的振荡或动作电位的一种规律的变化。
神经元发放的频率不高,但是震荡的频率可以很高(如在学习与记忆时)[33,34];可以从震荡的角度理解其与功能的关系。震荡有周期性,可以用来预测下一步的神经元活动,在同步震荡之中可以有信息传递的效果[35]。
在神经集群的水平上,大量神经元的同步活动可以产生宏观振荡,可以在脑电图中观察到。而发生在皮层尺度上的振荡便被称为脑电波。大尺度神经慢波震荡,对记忆固化有帮助,有多种传播模式,与结构传播有关系[36,37]。而不同回路中的波传播活动,可能是根据非线性作用产生的[38-42]。
神经系统是一个功能性的复杂系统,受长时间的进化压力,特别要考虑多种功能的优化,应该有一种成本的均衡。这种均衡应反映在功能-结构的关系上面[44]。大脑构建参数所消耗的能量大致运用在发放的产生及突触及信号的传播过程,还有神经元通讯之间神经递质的发放与重构之中[45]。也有团队进行了代谢与效益均衡的研究[46,47]。
对低成本-高效率的均衡的理解:从成本效益的角度理解生物神经系统,对于结构功能关系的生物物理基础及动力学机制来说是很重要的考虑视角,也是一个类脑计算的很好的参考。
通过限制网络结构组织,用经济性的连接方式,尽量的短,尽量的局部化,代谢上神经活动的发放率不应该很高,要稀疏,那如何有效地连接呢?可以参考文献[48-52]。
建模的目标
更好地理解数据(通过非参数处理如PCA(主成分分析)或参数化模型等方法)
探索隐藏在复杂数据下的工作原理和涌现原理
进行预测/帮助设计实验
建模的方案
明确建模的目标/尺度/元素(神经元,回路,大尺度,单个或多个尺度)
描述交互作用/复杂性的等级
决定模型的参数/通过数据校准
分析模型以理解机理/做出预测
建模的注意事项
(模型的)计算和分析复杂性vs对机制的理解
生物真实性vs数据可行性(缺失参数)
主要影响因子vs次要影响因子(物理学和生物学)
与数据结合的有现象性和机制性模型两种建模过程[53-55]。近年来也发展出一种介于现象性和机制性模型之间的基于机器学习的建模。
考虑到研究尺度的不同,单个神经元建模可以考虑神经元上的所有结构;大尺度或回路模型则将单个神经元简化,使用HH模型或者是Spike;发放率模型,更适合于大规模模拟;对于集群的模型,mass model用平均场模型描述集群的特性。
而多层次涌现现象则是单元特征与集群相互作用得到的新的特征,是大脑功能动力学的挑战。
单个神经元的发放如何与雪崩,震荡统一起来?这个统一有什么动力学机理?是否符合cost-efficient?
早在2003年便有研究人员提出基于电导的神经网络模型,在局部回路模型中,刺激简化为随机泊松过程输入,共有2500个神经元,80%为兴奋性神经元,考虑随机网络,突触耦合的定义为s(t)[56]。
实验中能观察到,神经系统出现震荡,但是神经元好几次震荡才会产生一次发放,这种模式下的神经元发放是无规则的[57]。
针对这种现象,有研究通过兴奋性,抑制性神经元网络进行建模:
I-I network[58]
E-I network[59]
Biological model: 兴奋和抑制因为相互作用,竞争关系而具有平衡态,耦合依赖于电压;Ising model: 耦合为电流,不依赖于电压[60]。
在本节介绍的基于电导的模型中也能观测到神经元活动现象:兴奋和抑制的电流耦合的紧密,当改变参数时,会出现中间自组织的情况。兴奋越来越快,抑制越来越慢时,同步越来越强。在生物合理的参数空间下同时涌现:
或是能观测到出现有部分关联但倾向于随机的过渡状态[61]。
从机理上,这处于一种兴奋-抑制的平衡态,并非平均值上的平衡,而是动态的平衡,网络较为稠密,而且兴奋-抑制连接有时间差,允许少部分兴奋性神经元发放,因网络是随机网络,神经元发放也是无规则的。模型也可以运用平均场理论近似简化为六维的模型。
神经元的发放可以表征信息。研究希望这样表征信息的熵比较高,通过对熵进行最大化推导,考虑信息的不同表征,可以得到不同表征结果[62,63]。
神经元的分布在空间并不均匀。随机网络是稀疏的,将其空间安排重新组织更贴近于模块化的生物模型,可以观察到有关联的神经发放,发放率下降,链接代价也下降, 而网络对外刺激相应更敏感, 实现“少即是多 (less is more)”[64]。
思考:扩展到更大的模型,这种模型如何处理信息,如何基于这种模型进行AI的开发?
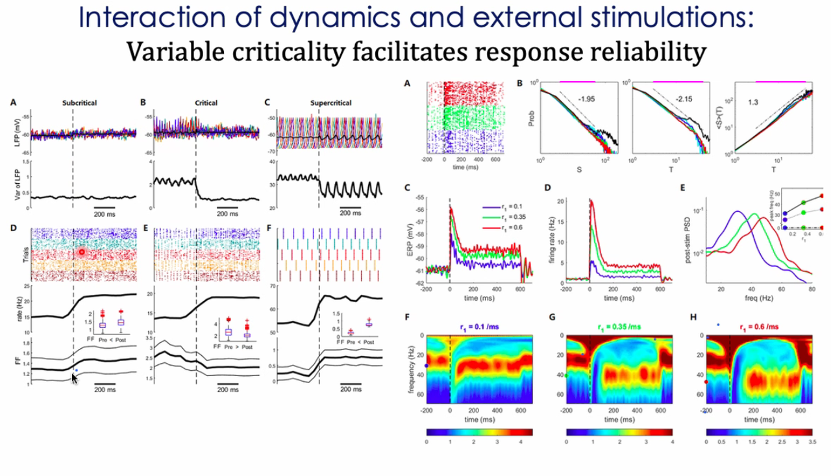
前面介绍过大脑有自发的活动,很多时候外界刺激要与神经活动交互。外界输入引起脑的相应的可变性(trail-to-trail variability)减小。很多不同尺度的例子证明静息态大脑处于临界态,即使在偏移的情况下也很快重回临界态。雪崩呈幂律分布。实验的亮度会改变gamma波的频率[65-69]。
通过研究实验看到的神经回路对外界响应的特点能否统一到模型中以及其与自组织临界态的模型的关系发现,图中A B C 比较不同的信号到来时间的trail-to-trail,信号到来前的可变性较大,信号到来后响应的可靠性变大。
平均场模型也能模拟这种情况。输入不改变临界态的稳定性,但是会改变可靠性[70]。
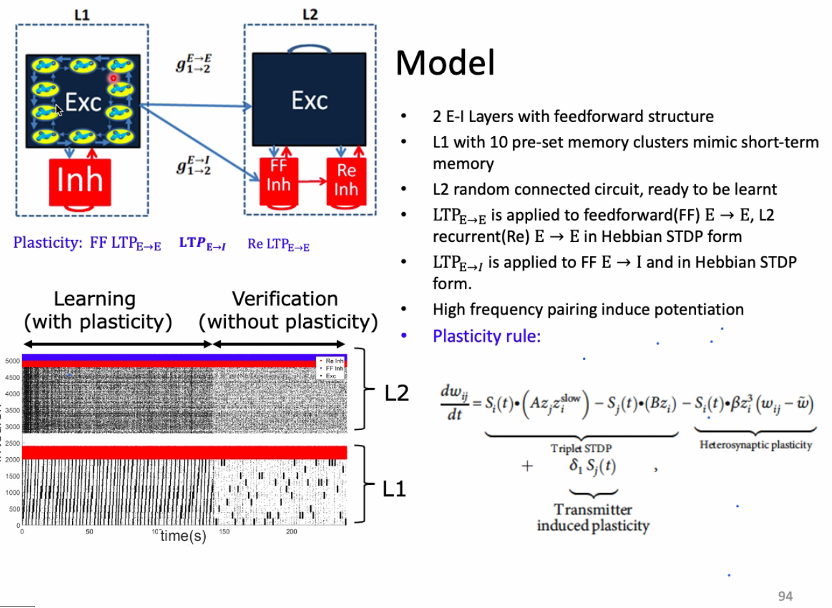
自发动力学对于神经系统突触的处理过程:如学习或记忆的相关研究。一组神经元构成一个集群来表征记忆。
与学习相关的记忆过程的可塑性:两个神经元之间的连接依赖于发放的时间,突触前先发放,受到驱动的神经元再发放,则连接加强。这种时间尺度的精度为毫秒级,新的实验发现这种突触可塑性也会影响其他神经元的连接。
学习与记忆过程中经常观测到gamma波,或是gamma和theta的整合。这些复杂的动力学如何通过可塑性影响学习与记忆?模型将学习记忆的动力学和网络连接共变化的环状机制分开成两部分进行研究,分别理解其中一个的作用,最后再整合起来研究共演化如何影响学习与记忆[71-73]。
在学习过程中,网络自组织共演化网络中,有外界驱动的神经元的连接会有很快地加强。学习的过程中能观测到神经元spike的模式变化很大,震荡中的gamma波的出现意味着学习效率的增长[74]。
小鼠E-I神经元突触有可塑性,受到γCaMKII蛋白酶影响,若将这种蛋白酶敲除,E-I可塑性消失,但E-E可塑性不受影响。并发现该操作不会影响短期记忆,但是将其固化为长期记忆便会受到此操作影响[75]。
图3. 行为方式与分析动力学的结合模型,左上角:左图为分子动力学影响,右图为行为方式。可以通过右下角模型将两者结合。
通过实验验证上述模型。实验上可能不易进行跨尺度研究,但建模会给出帮助并指导实验[76]。
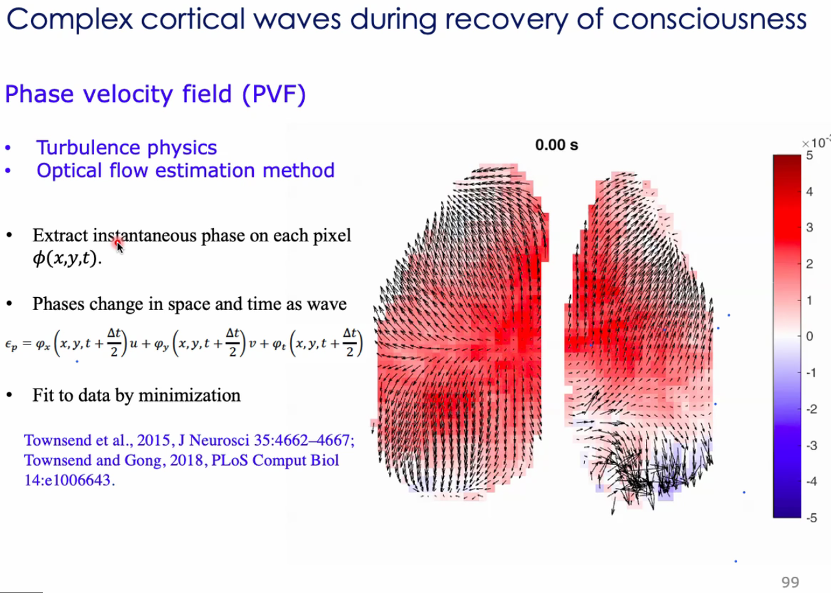
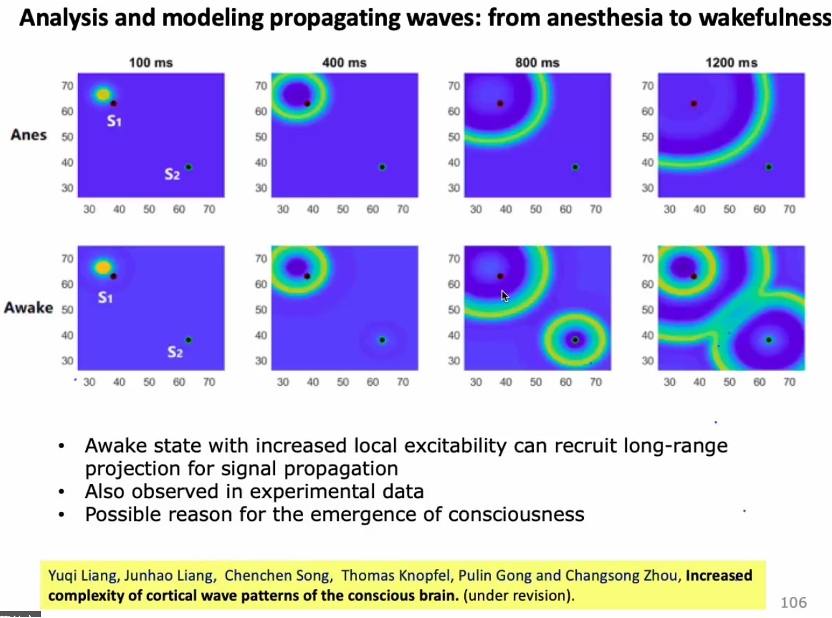
在分析波的传播时,用相速度场 (PVF) 刻画大脑点活动。时间、空间上波的传播与相位速度的变化有关,右图为麻醉情况下相位场变化动图[77,78]。
波源波汇出现位置与脑的大尺度解剖有关。信息处理在大脑中有层次性,如视觉信号在大脑中传播是从低级到高级脑区传播。按照这种排序,波源波汇更多出现在低级脑区[79]。
在考虑了空间传播后,有研究提出Neural field model(波动方程)。麻醉会影响抑制神经元的时间参数,改变脑区耦合的扩散强度,意为既改变了局部动力学,又改变了传输的效果。清醒时又会偶尔发现传播快的大型波动,模型与实验符合[80,81]。
从麻醉到清醒的动力学过程也是意识恢复的过程。通过模型可以理解局部回路在清醒时是兴奋性增强,这种局部变化会影响long range连接(脑区之间的连接)对信号传播的作用。如在麻醉时,对S1刺激,会逐渐传播到S2,但并不会引起S2的响应,因S2兴奋性不够大。清醒时long range能传播到S2形成新的传播源,与S1开始竞争。模型能够帮助理解清醒时大脑对外界响应,底层结构起作用,但是在麻醉时可能起不了很强的作用。
脑网络有什么特殊的结构特征?结构-功能关系又是如何?作为一个信息系统如何达到对信息的特异处理及整合?数据已有(动物的)结构网络和(人的)功能网络,但是人的结构网络测量(DTI)仍是较晚才出现[82-88]。
猫的脑网络中,各种子系统之间互有连接,称为hyper-community[89]。
猴子的脑区:网络脑区功能上是分开的,结构上是成块的。研究发现区域上分开的局部网络需要long range连接,少数脑区负责长程连接,而这些少数脑区内部连接十分紧密。这种集中对于信息整合很重要,对于功能分划也重要[90]。
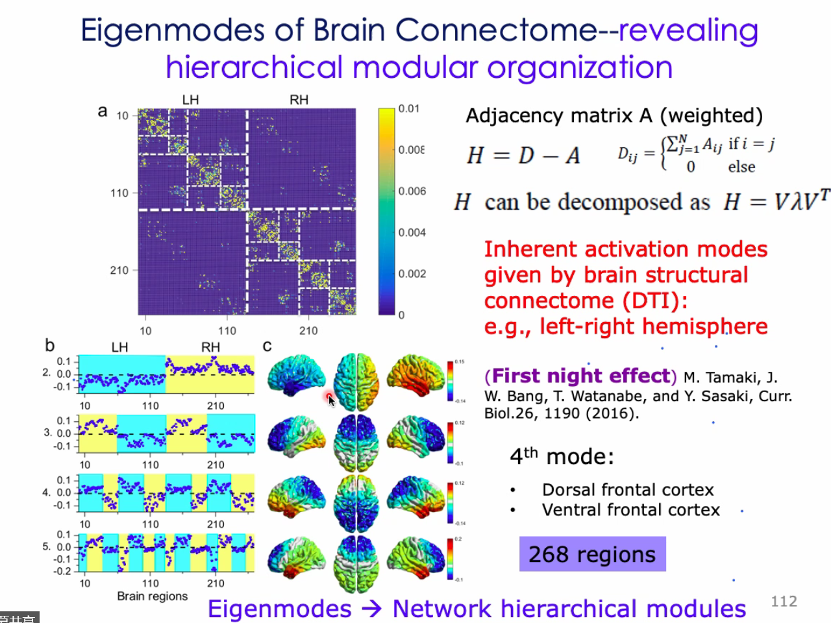
结构与动力学的关系方面:物理科学中运用晶体物理研究该关系:如原子间耦合关系的震动modes与声学、热学结构相关。通过邻接矩阵得到Laplacian矩阵,再进行特征向量分解,特征向量能够表征运动方向。给定一个随机的初始条件,则可以了解物体接下来的运动模式[91-95]。
对结构多层次模块的组织:DTI得到的结构性连接。 发现mode2正负分别对应左右脑,可能提供first night effect的一个结构基础。mode3又会细分地对应前后脑。将大脑分成268个脑区。
缺省模式(default mode):DMN能被分成前脑后脑或左右脑,在认知过程中可能会被单独地激发一部分[97-98]。
运用了很多神经元代表脑区的模型过于细致不利于分析,如下研究提出的线性模型可以直接写出脑区之间的功能性连接。给定耦合强度和结构连接(SC)可以直接分析出FC。可以再与empirical FC(如从BOLD序列得到的FC)作比较[99-101]。
但是将半脑内深层次连接打乱,其他不变,发现大脑网络的复杂性降低,完全随机化也能发现复杂性降低[102]。
进一步分析复杂性的组织形式:多层次模块划分的方法运用于功能连接(特征向量分解等)。如第一个mode脑区一致,数据中能量占比最大。可以不断地细分mode,得到不同level mode的能量占比[103]。
大脑在功能的整合-分离状态间一直切换[104]。研究运用高斯线性模型,发现耦合增加,功能的分离变弱,整合变强。正常人大脑平均来说处于分离-整合的平衡状态,但是个体会有差异。这种差异是否与个人能力有关[105]?人群中,不同脑区的变化与能力的变化是一致的(基于个体差异的建模)[106]。脑区之间也是不均匀的:不同脑区之间也有层次性,体现在神经递质的浓度,神经元密度等。那么如何把微观的神经解剖特性融合到大尺度模型?不同尺度的回路如何建模[107-114]?
因此对于大尺度结构功能网络:静息状态下,动力学组织能进行分离和整合,但是有个体差异;该分离和整合的差异与能力有关。
传统的构建神经网络模型往往需要从结构原理出发自下而上进行。而利用机器学习模型作为大脑模型,更像是从功能出发。一方面,人工学习出来的网络比大脑简单许多,但是完成任务较好。另一方面,通过训练模型,人工神经网络可以启发我们开发与人类智能相关的AI。如考虑E-I平衡也可以作为正则化条件放入训练[115]。
多任务态训练揭示了神经活动规则:不同的外界信号输入大脑,对应不同的神经活动对任务进行表征,表现出大脑多尺度模块化的组织[116,117]。
时间处理的建模理念(Dr. Bi Zedong, Institute for Future Qingdao University):工作记忆如何处理时间?空间信息如何表征?网络中什么结构可以支撑时间的表征[118]?
通过训练(刺激),得到动作神经元在时间限内给出反应。在工作记忆中,脑的状态会变化,趋于一个吸引子或瞬态(transient)[119,120]。
在对大脑的研究过程中,我们会发现很多问题:数据复杂,对于数据之下的组织机制不理解,或是不能想象数据产生的过程,很难想象这种无规则的发放在空间中体现出一种组织现象。如何应对数据上的诸多问题?也许我们需要从数学建模的角度理解隐藏在数据背后的神经活动机制。在动力学下刻画大脑处理信息的机制[121-137]。
一般来说,神经动力学作为一个跨学科的研究,构建复杂脑动力学和认知,非线性动力学的方法与心理学,神经科学的合作,可以研究更多更具体的问题。未来的神经科学研究中,可能更会出现这样的合作关系:神经科学(生物实验)– 数据分析抽取特征-建模和理论分析、数据处理与机制研究。
生物的智能可能就隐藏在大脑的复杂性之中。如何理解这种复杂性?理解智能的表征?这仍是一个挑战[138]。
[1] Sejnowski T., Churchland P., Movshon J.. Putting big data to good use in neuroscience. Nat Neurosci, 2014, 17: 1440–1441
[2] J Hellyer P., M Shanahan, G Scott. The control of global brain dynamics: opposing actions of frontoparietal control and default mode networks on attention[J]. Journal of Neuroscience, 2014, 34(2): 451–461
[3] Garrett. Moment-to-moment brain signal variability: A next frontier in human brain mapping?. Neuroscience and Biobehavioral Reviews, 2013
[4] Gómez C., Poza J., Fernández A.. Entropy analysis of MEG background activity in attention-deficit/hyperactivity disorder[C]//2013. 35th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC, 2013: 5057–5060
[5] O Sokunbi M., W Fung, V Sawlani. Resting state fMRI entropy probes complexity of brain activity in adults with ADHD[J]. Psychiatry Research: Neuroimaging, 2013, 214(3): 341–348
[6] A Méndez M., P Zuluaga, R Hornero. Complexity analysis of spontaneous brain activity: effects of depression and antidepressant treatment[J]. Journal of psychopharmacology, 2012, 26(5): 636–643
[7] Takahashi T., Y Cho R., Mizuno T.. Antipsychotics reverse abnormal EEG complexity in drug-naive schizophrenia: a multiscale entropy analysis[J]. Neuroimage, 2010, 51(1): 173–182
[8] R A., N Kovačević, R McIntosh A.. Brain signal variability relates to stability of behavior after recovery from diffuse brain injury[J]. Neuroimage, 2012, 60(2): 1528–1537
[9] Levy R.B., Reyes A.D.. Spatial profile of excitatory and inhibitory synaptic connectivity in mouse primary auditory cortex. J. Neurosci, 2012, 32: 5609–5619
[10] Ercsey-Ravasz M.. A predictive network model of cerebral cortical connectivity based on a distance rule. Neuron, 2013, 80(16): 184–197
[11] John M. Beggs, Nicholas Timme. Critical brain networks. Front. Physiol., 2012
[12] Softky W.R., Koch C.. The highly irregular firing of cortical cells is inconsistent with temporal integration of random EPSPs. J Neurosci, 1993, 13: 334–350
[13] Shadlen M.N., Newsome W.T.. The variable discharge of cortical neurons: implications for connectivity, computation, and information coding. The Journal of neuroscience: the official journal of the Society for Neuroscience, 1998, 18(10): 3870–96
[14] Shu, Yousheng, Hasenstaub, et al. Turning on and off recurrent balanced cortical activity. nature, 2003, 423(6937): 288-293
[15] Froemke R., Merzenich M., Schreiner C.. A synaptic memory trace for cortical receptive field plasticity. Nature, 2007, 450: 425–429
[16] van Vreeswijk C., Sompolinsky H.. Chaos in neuronal networks with balanced excitatory and inhibitory activity. Science, 1996, 274: 1724–1726
[17] Okun M., Lampl I.. Instantaneous correlation of excitation and inhibition during ongoing and sensory-evoked activities[J]. Nature neuroscience, 2008, 11(5): 535–537
[18] Elad Schneidman, Michael J. Berry II, Ronen Segev & William Bialek. Weak pairwise correlations imply strongly correlated network states in a neural population. nature, 2006
[19] Friedman N.. Universal critical dynamics in high resolution neuronal avalanche data. Phys. Rev. Lett, 2012, 108: 208102
[20] Fontenele A.J., de Vasconcelos N.A.P., Feliciano T., et al. Criticality between cortical statesPhys. Rev. Lett, 2019, 122: 208101
[21] Beggs J.M., Plenz D.. Neuronal avalanches in neocortical circuits. J. Neurosci, 2003, 23: 11167–11177
[22] E Gireesh, D Plenz. Neuronal avalanches organize as nested theta- and beta/gamma-oscillations during development of cortical layer 2/3. Proc Natl Acad Sci USA, 2008, 105: 7576–7581,
[23] Petermann T.. Spontaneous cortical activity in awake monkeys composed of neuronal avalanches. Proc. Natl Acad. Sci. USA, 2009, 106: 15921–15926
[24] Scott G., D Fagerholm E., Mutoh H.. Voltage imaging of waking mouse cortex reveals emergence of critical neuronal dynamics[J]. Journal of Neuroscience, 2014, 34(50): 16611–16620
[25] D Fagerholm E., G Scott, L Shew W.. Cortical entropy, mutual information and scale-free dynamics in waking mice[J]. Cerebral cortex, 2016, 26(10): 3945–3952
[26] L Shew W., P Clawson W., J Pobst. Adaptation to sensory input tunes visual cortex to criticality[J]. Nature Physics, 2015, 11(8): 659–663
[27] Zhang D., Raichle M.E.. Disease and the brain’s dark energy. Nat. Rev. Neurol, 2010, 6: 15–28
[28] Tagliazucchi E., Balenzuela P., Fraiman D.. Criticality in large-scale brain fMRI dynamics unveiled by a novel point process analysis[J]. Frontiers in physiology, 2012, 3: 15
[29] Luca Cocchi,Leonardo L. Gollo,Andrew Zalesky,Michael Breakspear. Criticality in the brain: A synthesis of neurobiology, models and cognition. Progress in neurobiology, 2017, 158: 132-152
[30] Shew W.L., Yang H., Petermann T., et al. Neuronal avalanches imply maximum dynamic range in cortical networks at criticality. J. Neurosci, 2009, 29: 15595–15600
[31] Shew W.L., Yang H., Yu S., et al. Information capacity and transmission are maximized in balanced cortical networks with neuronal avalanches. J. Neurosci, 2011, 31: 55–63
[32] Yang H., Shew W.L., Roy R., et al. PlenzMaximal variability of phase synchrony in cortical networks with neuronal avalanches. Journal of Neuroscience, 2012, 32: 1061–1072
[33] Linkenkaer-Hansen K., Nikouline V.V., Palva J.M., et al. Long-range temporal correlations and scaling behavior in human brain oscillations. J. Neurosci, 2001, 21: 1370–1377
[34] Fisahn A., Pike F.G., Buhl E.H., et al. Cholinergic induction of network oscillations at 40 Hz in the hippocampus in vitro. Nature, 1998, 394: 186–189
[35] Buzsaki G., Draguhn A.. Neuronal oscillations in cortical networks[J]. science, 2004, 304(5679): 1926–1929
[36] Massimini M., Huber R., Ferrarelli F.. The sleep slow oscillation as a traveling wave[J]. Journal of Neuroscience, 2004, 24(31): 6862–6870
[37] Luczak A., Barthó P., L Marguet S.. Sequential structure of neocortical spontaneous activity in vivo[J]. Proceedings of the National Academy of Sciences, 2007, 104(1): 347–352
[38] R Xulvi-Brunet, I M Sokolov. Reshuffling scale-free networks: From random to assortative. Physical Review E, 2005
[39] Wang S.J., Ouyang G., Guang J., et al. Stochastic Oscillation in Self-Organized Critical States of Small Systems: Sensitive Resting State in Neural Systems. Phys Rev Lett, 2016, 116(1)
[40] K Sato T., I Nauhaus, M Carandini. Traveling waves in visual cortex[J]. Neuron, 2012, 75(2): 218–229
[41] Muller L., Chavane F., Reynolds J.. Cortical travelling waves: mechanisms and computational principles[J]. Nature Reviews Neuroscience, 2018, 19(5): 255–268
[42] Nir Y.. Regional slow waves and spindles in human sleep. Neuron, 2011, 70: 153–169
[43] Shimaoka D., Song C., Knöpfel T.. State-dependent modulation of slow wave motifs towards awakening[J]. Frontiers in cellular neuroscience, 2017, 11: 108
[44] Merolla P.A., Arthur J.V., Alvarez-Icaza R., et al. A million spiking-neuron integrated circuit with a scalable communication network and interface. Science, 2014, 345(6197): 668–673
[45] Laughlin S.B.. Energy as a constraint on the coding and processing of sensory information. Curr. Opin. Neurobiol, 2001, 11,475-480
[46] G Vlassenko A., N Vaishnavi S., L Couture. Spatial correlation between brain aerobic glycolysis and amyloid-β (Aβ) deposition[J]. Proceedings of the National Academy of Sciences, 2010, 107(41): 17763–17767
[47] Chen Yuhan, Zhang Zi-Ke, He Yong, et al. A Large-Scale High-Density Weighted Structural Connectome of the Macaque Brain Acquired by Predicting Missing Links. proceedings of the national academy of sciences, 2020, 30(1): 4771–4789
[48] Chen Y., Wang S., C Hilgetag C.. Trade-off between multiple constraints enables simultaneous formation of modules and hubs in neural systems[J]. PLoS computational biology, 2013, 9(3): 1002937
[49] Chen Y.. Features of spatial and functional segregation and integration of the primate connectome revealed by trade-off between wiring cost and efficiency. PLOS Comput. Biol, 2017, 13: 1005776
[50] Chen Yuhan, Zhang Zi-Ke, He Yong, et al. A Large-Scale High-Density Weighted Structural Connectome of the Macaque Brain Acquired by Predicting Missing Links. proceedings of the national academy of sciences, 2020, 30(1): 4771–4789
[51] P Yang D., J Zhou H., C Zhou. Co-emergence of multi-scale cortical activities of irregular firing, oscillations and avalanches achieves cost-efficient information capacity[J]. PLoS computational biology, 2017, 13(2): 1005384
[52] Liang J., Zhou T., Zhou C.. Hopf bifurcation in mean field explains critical avalanches in excitation-inhibition balanced neuronal networks: a mechanism for multiscale variability[J]. Frontiers in systems neuroscience, 2020: 87
[53] Huang C.. Circuit models of low-dimensional shared variability in cortical networks. Neuron, 2019, 101: 337–348
[54] Goris R.L.T., Movshon J.A., Simoncelli E.P.. Partitioning neuronal variability. Nat. Neurosci, 2014, 17: 858–865
[55] Lin I.-C., Okun M., Carandini M., et al. The nature of shared cortical variability. Neuron, 2015, 87: 644–656
[56] Brunel N., Wang X.-J.. What determines the frequency of fast network oscillations with irregular neural discharges? I. Synaptic dynamics and excitation-inhibition balance. J. Neurophysiol, 2003, 90: 415–430
[57] N Brunel, V Hakim. Sparsely synchronized neuronal oscillations. Chaos, 2008, 18: 15–113,
[58] Brunel N., Hakim V.. Fast global oscillations in networks of integrate-and-fire neurons with low firing rates. Neural Comput, 1999, 11: 1621–1671
[59] Brunel N.. Dynamics of sparsely connected networks of excitatory and inhibitory spiking neurons. Journal of Computational Neuroscience, 2000, 8(3): 183–208
[60] van Vreeswijk C., Sompolinsky H.. Chaos in neuronal networks with balanced excitatory and inhibitory activity. Science, 1996, 274: 1724–1726
[61] P Yang D., J Zhou H., C Zhou. Co-emergence of multi-scale cortical activities of irregular firing, oscillations and avalanches achieves cost-efficient information capacity[J]. PLoS computational biology, 2017, 13(2): 1005384
[62] Laughlin S.B.. Energy as a constraint on the coding and processing of sensory information. Curr. Opin. Neurobiol, 2001, 11,475-480
[63] P Yang D., J Zhou H., C Zhou. Co-emergence of multi-scale cortical activities of irregular firing, oscillations and avalanches achieves cost-efficient information capacity[J]. PLoS computational biology, 2017, 13(2): 1005384
[64] Sheng-Jun Wang, Junhao Liang, Changsong Zhou. Less is More: Wiring-Economical Modular Networks Support Self-Sustained Firing-Economical Neural Avalanches for Efficient Processing. arXiv:2007.02511, 2020
[65] Churchland M., Yu B., Cunningham J.. Stimulus onset quenches neural variability: a widespread cortical phenomenon, 2010, 13: 369–378
[66] He B.J.. Spontaneous and task-evoked brain activity negatively interact, 2013, 33(11): 4672–4682
[67] L Shew W., P Clawson W., J Pobst. Adaptation to sensory input tunes visual cortex to criticality[J]. Nature Physics, 2015, 11(8): 659–663
[68] Ray S., Maunsell J.H.. Differences in gamma frequencies across visual cortex restrict their possible use in computation, 2010, 67: 885–896
[69] Ray S., Maunsell J.H.R.. Do gamma oscillations play a role in cerebral cortex?, 2015, 19: 78–85
[70] Liang J., Zhou C.. Criticality enhances the multilevel reliability of stimulus responses in cortical neural networks[J]. PLoS computational biology, 2022, 18(1): 1009848
[71] Ray S., Maunsell J.H.R.. Do gamma oscillations play a role in cerebral cortex?, 2015, 19: 78–85
[72] Dan Y., Poo M.-M.. Spike timing-dependent plasticity of neural circuits, 2004, 44: 23–30
[73] Zenke F.. Diverse synaptic plasticity mechanisms orchestrated to form and retrieve memories in spiking neural networks, 2015, 6(6922)
[74] Li, Kwan Tung, Liang, et al. Gamma Oscillations Facilitate Effective Learning in Excitatory-Inhibitory Balanced Neural Circuits. neural plasticity, 2021, 2021
[75] X. He, J. Li, G. Zhou, et al. Gating of hippocampal rhythms and memory by synaptic plasticity in inhibitory interneurons, 2021, 109: 1013–1028 10 1016
[76] Li Kwan Tung, HE Xingzhi, Zhou Guangjun, et al. Rational designing of oscillatory rhythmicity for memory rescue in plasticity-impaired learning networks, Cell Reports
[77] Townsend R.G.. Emergence of complex wave patterns in primate cerebral cortex, 2015, 35: 4657–4662
[78] Townsend R.G., Gong P.. Detection and analysis of spatiotemporal patterns in brain activity, 2018, 14(12): 1006643
[79] Liang Y., Song C., Liu M.. Cortex-wide dynamics of intrinsic electrical activities: propagating waves and their interactions[J]. Journal of Neuroscience, 2021, 41(16): 3665–3678
[80] Steyn-Ross M.L., Steyn-Ross D.A., Sleigh J.W.. Interacting turing-Hopf instabilities drive symmetry-breaking transitions in a mean-field model of the cortex: a mechanism for the slow oscillation, 2013, X: 3
[81] Rudolph U., Antkowiak B.. Molecular and neuronal substrates for general anaesthetics, 2004, 5: 709–720
[82] Sporns O., Chialvo D., Kaiser M., et al. Organization, development and function of complex brain networks, 2004, 8: 418–425
[83] Zhou C., Zemanova L., Zamora G., et al. Hierarchical organization unveiled by functional connectivity in complex brain networks, 2006, 97: 238103
[84] Zemanová L., Zhou C., Kurths J.. Structural and functional clusters of complex brain networks, 2006, 224: 202–212
[85] Wang Z., Dai Z., Gong G., et al. Understanding structural-functional relationships in the human brain: A large-scale network perspective, 2015
[86] Zamora-Lopez G., Zhou C., Kurths J.. Cortical hubs form a module for multisensory integration on top of the hierarchy of cortical networks, 2010, 4, 1
[87] Zhao M., Zhou C., Lü J., et al. Competition between intra-community and inter-community synchronization and relevance in brain cortical networks, 2011, 84(1): 016109
[88] Zhou C., Zemanová L., Zamora-López G., et al. Structure–function relationship in complex brain networks expressed by hierarchical synchronization, 2007, 9: 178
[89] Chen Y.. Features of spatial and functional segregation and integration of the primate connectome revealed by trade-off between wiring cost and efficiency. PLOS Comput. Biol, 2017, 13: 1005776
[90] Raj A.. A network diffusion model of disease progression in dementia, 2012, 73: 1204–1215
[91] F. Abdelnour, U. Voss H., A Raj. Network diffusion accurately models the relationship between structural and functional brain connectivity networks, 2014, 90: 335–347
[92] Atasoy S., Donnelly I., Pearson J.. Human brain networks function in connectome-specific harmonic waves, 2016, 7: 10340
[93] Wang Maxwell B., Owen Julia P., Mukherjee Pratik, et al. Brain network eigenmodes provide a robust and compact representation of the structural connectome in health and disease, 2017, 13(6): 1–20
[94] Abdelnour F.. Functional brain connectivity is predictable from anatomic network’s Laplacian eigen-structure, 2018, 172: 728–739
[95] Tamaki M.. Night watch in one brain hemisphere during sleep associated with the first-night effect in humans, 2016, 9: 1190–1194
[96] Greicius M.D., Krasnow B., Reiss A.L., et al. Functional connectivity in the resting brain: a network analysis of the default mode hypothesis, 2003, 100: 253–258
[97] Wu X.J.. Functional network connectivity alterations in schizophrenia and depression, 2017, 263: 113–120
[98] Tononi G., Sporns O., Edelman G.M.. A measure for brain complexity: relating functional segregation and integration in the nervous system, 1994, 91: 5033–5037
[99] Zamora-Lopez G., Zhou C., Kurths J.. Exploring brain function from anatomical connectivity, 2011, 5: 83
[100] Zamora-López G., Chen Y., Deco G.. Functional complexity emerging from anatomical constraints in the brain: the significance of network modularity and rich-clubs[J]. Scientific reports, 2016, 6(1): 1–18
[101] Rong Wang, Pan Lin, Mianxin Liu, et al. Hierarchical Connectome Modes and Critical State Jointly Maximize Human Brain Functional Diversity. physical review letters, 2019, 123(3): 038301
[102] Rong Wang, Mianxin Liu, Xinhong Cheng, et al. Segregation, integration, and balance of large-scale resting brain networks configure different cognitive abilities. proceedings of the national academy of sciences, 2021, 118(23)
[103] Rong Wang, Mianxin Liu, Xinhong Cheng, et al. Segregation, integration, and balance of large-scale resting brain networks configure different cognitive abilities. proceedings of the national academy of sciences, 2021, 118(23)
[104] Rong Wang, Mianxin Liu, Xinhong Cheng, et al. Segregation, integration, and balance of large-scale resting brain networks configure different cognitive abilities. proceedings of the national academy of sciences, 2021, 118(23)
[105] Kristanto D., Liu X., Sommer W., et al. What do anatomical properties of the brain reveal about the ontology of human cognitive abilities?. arXiv:2108.00466, 2021
[106] Rishidev Chaudhuri. A Large-Scale Circuit Mechanism for Hierarchical Dynamical Processing in the Primate Cortex. neuron, 2015, 88(2): 419-431
[107] Joglekar M.R., Mejias J.F., Yang G.R., et al. Inter-areal Balanced Amplification Enhances Signal Propagation in a Large-Scale Circuit Model of the Primate Cortex, 2018, 98: 222–234 8
[108] Wang P.. Inversion of a large-scale circuit model reveals a cortical hierarchy in the dynamic resting human brain, 2019, 5: 7854
[109] Demirtaş M.. Hierarchical heterogeneity across human cortex shapes large-scale neural dynamics, 2019, 101: 1181–1194 13
[110] Deco G., Kringelbach M.L., Arnatkeviciute A., et al. Dynamical consequences of regional heterogeneity in the brain’s transcriptional landscape, 2021, 7(29): 4752
[111] Zhou C., Zemanova L., Zamora G., et al. Hierarchical organization unveiled by functional connectivity in complex brain networks, 2006, 97: 238103
[112] Honey C.J., Kötter R., Breakspear M., et al. Network structure of cerebral cortex shapes functional connectivity on multiple time scales, 2007, 104: 10240–10245
[113] Rong Wang, Pan Lin, Mianxin Liu, et al. Hierarchical Connectome Modes and Critical State Jointly Maximize Human Brain Functional Diversity. physical review letters, 2019, 123(3): 038301
[114] Song H.F., Yang G.R., Wang X.J.. Training excitatory-inhibitory recurrent neural networks for cognitive tasks: a simple and flexible framework, 2016, 12(2): 1004792
[115] Yang G.R., Joglekar M.R., Song H.F.. Task representations in neural networks trained to perform many cognitive tasks. Nat Neurosci, 2019, 22: 297–306
[116] Yang G.R., Wang X.J.. Artificial neural networks for neuroscientists: A primer, 2020, 107(6): 1048–1070
[117] Bi Z., Zhou C.. Understanding the computation of time using neural network models[J]. Proceedings of the National Academy of Sciences, 2020, 117(19): 10530–10540
[118] Jin D.Z., Fujii N., Graybiel A.M.. Neural representation of time in cortico-basal ganglia circuits, 2009, 106(45): 19156–19161
[119] Mita A., Mushiake H., Shima K., et al. Interval time coding by neurons in the presupplementary and supplementary motor areas, 2009, 12(4): 502–507
[120] Sheng-Jun Wang, Junhao Liang, Changsong Zhou. Less is More: Wiring-Economical Modular Networks Support Self-Sustained Firing-Economical Neural Avalanches for Efficient Processing. arXiv:2007.02511, 2020
[121] Rong Wang, Mianxin Liu, Xinhong Cheng, et al. Segregation, integration, and balance of large-scale resting brain networks configure different cognitive abilities. proceedings of the national academy of sciences, 2021, 118(23)
[122] Liang Y., Song C., Liu M.. Cortex-wide dynamics of intrinsic electrical activities: propagating waves and their interactions[J]. Journal of Neuroscience, 2021, 41(16): 3665–3678
[123] Chen Yuhan, Zhang Zi-Ke, He Yong, et al. A Large-Scale High-Density Weighted Structural Connectome of the Macaque Brain Acquired by Predicting Missing Links. proceedings of the national academy of sciences, 2020, 30(1): 4771–4789
[124] Bi Z., Zhou C.. Understanding the computation of time using neural network models[J]. Proceedings of the National Academy of Sciences, 2020, 117(19): 10530–10540
[125] Yuhan Chen, Zi-Ke Zhang, Yong He, et al. A large-scale high-density weighted structural connectome of the macaque brain acquired by predicting missing links. Cerebral Cortex, 2020, 30(9): 4771–4789
[126] Liu M., Liu X., Hildebrandt A., et al. Individual cortical entropy profile: Test–retest reliability, predictive power for cognitive ability, and neuroanatomical foundation, 2020, 1(1): 015
[127] Kristanto D., Liu M., Liu X., et al. Predicting reading ability from brain anatomy and function: From areas to connections, 2020, 218: 116966
[128] Rong Wang, Pan Lin, Mianxin Liu, et al. Hierarchical Connectome Modes and Critical State Jointly Maximize Human Brain Functional Diversity. physical review letters, 2019, 123(3): 038301
[129] Liu X., Hildebrandt A., Meyer K., et al. Patterns of individual differences in fiber tract integrity of the face processing brain network support neurofunctional models, 2020, 204: 116229
[130] Liu M., Song C., Liang Y., et al. Assessing spatiotemporal variability of brain spontaneous activity by multiscale entropy and functional connectivity, 2019, 198: 198–220
[131] Ouyang G., Hildebrandt A., Sommer W., et al. Exploiting the intra-subject latency variability from single-trial event-related potentials in the P3 time range: a review and comparative evaluation of methods, 2017, 75: 1–21
[132] Chen Y.. Features of spatial and functional segregation and integration of the primate connectome revealed by trade-off between wiring cost and efficiency. PLOS Comput. Biol, 2017, 13: 1005776
[133] P Yang D., J Zhou H., C Zhou. Co-emergence of multi-scale cortical activities of irregular firing, oscillations and avalanches achieves cost-efficient information capacity[J]. PLoS computational biology, 2017, 13(2): 1005384
[134] Wang S.J., Ouyang G., Guang J., et al. Stochastic Oscillation in Self-Organized Critical States of Small Systems: Sensitive Resting State in Neural Systems. Phys Rev Lett, 2016, 116(1)
[135] Chen Y., Wang S., C Hilgetag C.. Trade-off between multiple constraints enables simultaneous formation of modules and hubs in neural systems[J]. PLoS computational biology, 2013, 9(3): 1002937
[136] Ouyang G., Schacht A., Zhou C., et al. Overcoming limitations of the ERP method with R esidue I teration D ecomposition (RIDE): A demonstration in go/no‐go experiments, 2013, 50(3): 253–265
[137] Zhou C., Zemanova L., Zamora G., et al. Hierarchical organization unveiled by functional connectivity in complex brain networks, 2006, 97: 238103
[138]Benjamin Cramer, David Stöckel, Markus Kreft, et al. Control of criticality and computation in spiking neuromorphic networks with plasticity. nature communications, 2020, 11(1): 1-11
随着电生理学、网络建模、机器学习、统计物理、类脑计算等多种技术方法的发展,我们对大脑神经元相互作用机理与连接机制,对意识、语言、情绪、记忆、社交等功能的认识逐渐深入,大脑复杂系统的谜底正在被揭开。为了促进神经科学、系统科学、计算机科学等领域研究者的交流合作,我们发起了【神经动力学模型读书会】。
【神经动力学模型读书会】围绕神经网络多尺度建模及其在脑疾病、脑认知方面的应用进行了研讨。目前已经完结,读书会所形成的神经动力学科研社群集结了近500位相关领域的师生及产业界人士。现在报名读书会即可解锁录播视频并入群,欢迎从事相关研究、对相关领域有浓厚兴趣的探索者报名参与交流。
详情请见:
神经动力学模型读书会启动:整合计算神经科学的多学科方法