10月2日,2023年诺贝尔生理学或医学奖宣布授予 Katalin Karikó 和Drew Weissman,表彰他们在核苷碱基修饰方面的发现,这些发现帮助开发出有效的 mRNA 疫苗来对抗 COVID-19。mRNA 疫苗被认为是一场药物形式的革命,相关技术疗法具有广阔的应用前景,不仅能对抗传染病,还可以利用 mRNA 技术来定制癌症疫苗。
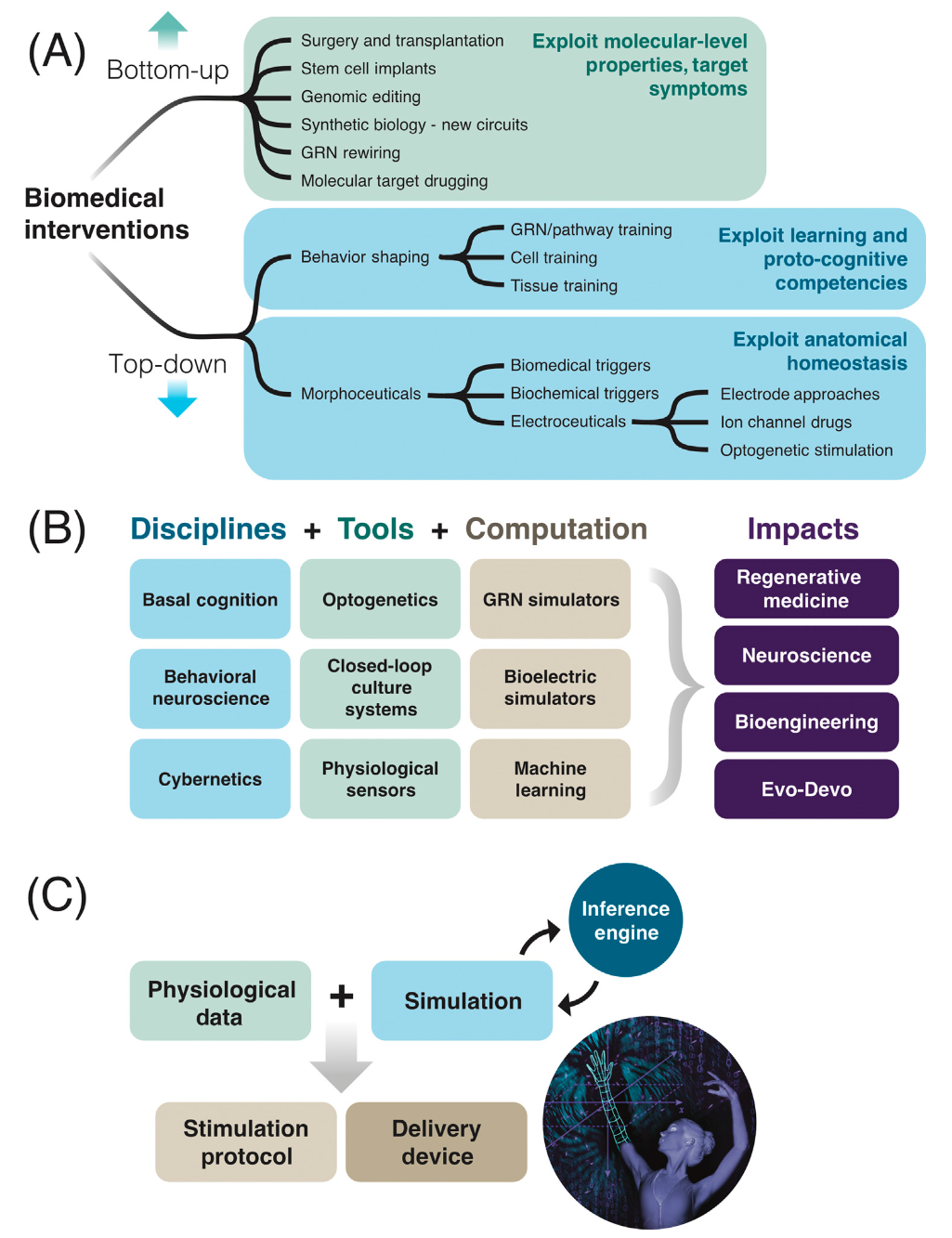
目前的生物医学方法主要是通过使用 CRISPR、基因治疗、蛋白工程等,改变微观基因产物,自下而上操纵细胞的“硬件”。从根本上而言,人体的复杂性并非直接由基因组指定,生物体以细胞为基本单元,形成各种集体行为,在从细胞到组织、器官、生物体和生态系统的各个层次上都表现出多样性和组织性,生命的“软件”具有模块化、自上而下的控制和多尺度能力等显著特征,使得生命具有强大的适应能力,但同时也为医学理解和医疗医药研发带来巨大的难题。
塔夫茨大学发育与合成生物学教授 Michael Levin 与匹兹堡大学病理学系副教授 Eric Lagasse 近期于 Cell 子刊 Trends in Molecular Medicine 发表综述文章“未来医学展望:从分子通道到集体智能”,指出生命形式的多尺度能力为生物医学提供了一条新的道路,利用组织和器官的固有集体智能。综述尤其突出利用生物电学和行为神经科学的进展,设计诱导结构和功能的自我修复方法,勾勒出通往“身心医学”的路线图。通过放松细胞控制机制是静态的假设,利用控制论、行为科学和发育生物学的强大概念,可能会为当前的生物医学挑战提供明确的解决方案。
研究领域:生命复杂性,生物医学,集体智能,神经科学
Eric Lagasse、Michael Levin | 作者
刘培源 | 译者

论文题目:
Future medicine: from molecular pathways to the collective intelligence of the body
论文地址:https://www.cell.com/trends/molecular-medicine/fulltext/S1471-4914(23)00142-9
二、身体智能:解剖稳态、集体智能和功能需求驱动的修复
三、“肝稳态”:细胞智能及其在肝病临床应用中的例证
四、利用身体智慧调节再生:神经生物学和行为科学指引方向
八、结论
复杂生物体所展示的显著解剖稳态表明,它们本质上是可重新编程的信息处理系统,提供了许多接口来解决其生理和解剖问题。这篇综述文章梳理了相关数据,表明生命形式的多尺度能力为生物医学提供了一条新的道路,利用组织和器官的固有集体智能。组织层面的自适应目标导向概念已经在临床实践中取得了成果。本文尤其突出利用生物电学和行为神经科学的进展,设计诱导结构和功能的自我修复方法,勾勒出通往“身心医学”的路线图。通过放松细胞控制机制是静态的假设,利用控制论、行为科学和发育生物学的强大概念,可能会为当前的生物医学挑战提供明确的解决方案。
到2060年,每年估计将有4800万人(全球死亡人数的47%)在生命的最后阶段因疾病和治疗而遭受巨大痛苦。在生命末期,为了修补老化的身体而进行越来越昂贵的干预措施,给医疗系统和整个社会带来了巨大负担。希望在于,认识到各种医疗需求要归结为一个关键能力:控制细胞集体所构建的解剖结构。如果我们了解细胞集体如何决定构建什么以及何时停止,我们就可以修复先天缺陷,再生因创伤性损伤、退行性疾病和衰老而失去的部分,并将癌细胞重新编程为正常组织。
人体表现的复杂性并非直接由基因组指定,而是由一组胚胎细胞的活动产生的(图1A)。这些细胞集体的计算和行为子程序对生长和形态做出解剖决策,而我们在修复或再生缺失或受损器官时必须针对并修改这些决策。身体是通过多尺度能力架构(图1B和框1)构建的——每个层次处理信息以解决生理、转录、解剖和行为空间的问题。学会利用它们的能力并管理细胞的感知[1]、记忆和设定点(不仅仅是当前的生化状态)为生物医学提供了一个强大的路线图,以实现持久的治愈。因此,全面再生医学的最终目标是能够将期望的组织和器官水平结果传达给细胞群体。
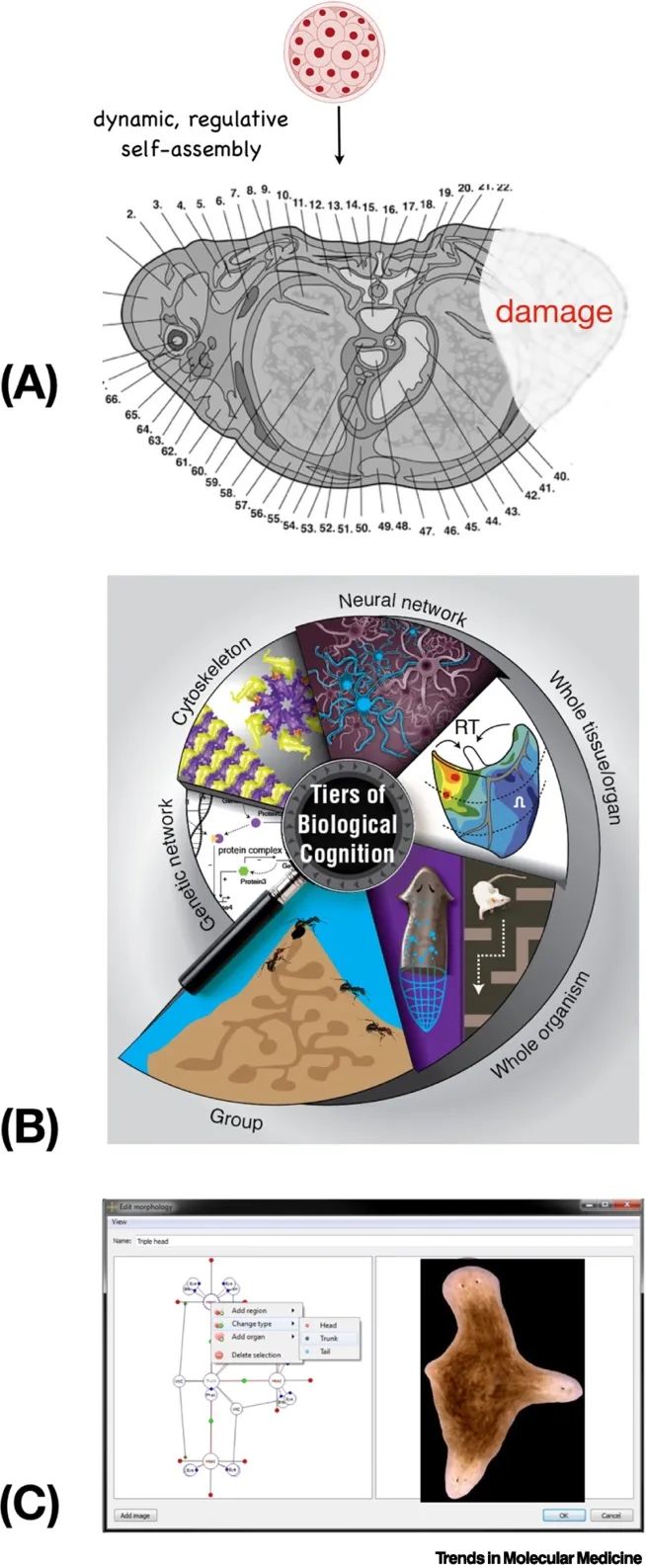
图1. 解剖编译器和多尺度能力。(A)人体的复杂性(在此以成年躯干的横截面显示)并非直接由基因组(编码亚细胞硬件-蛋白质)指定,而是由胚胎细胞团的活动产生的。(B)身体通过多尺度能力架构构建,其中每个层次处理信息以解决生理、转录、解剖和行为空间中的问题。(C)构建“解剖编译器”举例,用户能够指定任何解剖形状,并将其转换为一组刺激,必须提供给细胞以指导它们生长这种形状(如此处所示的三头扁虫)。解剖编译器不会是一个3D打印机或用于微观管理基因表达或干细胞命运的设备——实际上它将是一个将用户的解剖目标转换为细胞集体中目标形态信息的重新规范的通信设备。
这个目标可以概括为以下设计挑战:创建一个解剖编译器——一种计算机软件,它接受任何所需结构(生物器官或附属物,甚至是新颖的合成形态)的解剖规范,并输出激励细胞构建完全相同结构所需的信号列表。对生物医学和合成生物工程来说,对生长和形态具有完全控制力是至关重要的目标。关键是,解剖编译器不是一个3D打印机或其他微观管理细胞生长结构和功能的方式;在重要意义上,它是一个与细胞群体的集体智能进行交互的通信设备,是人类生物医学目标与形态发生稳态、异稳态和同稳态设定点之间的翻译者(图1C)。这代表了解决当前生物医学限制的根本性新方法。
药物发现对于复杂疾病状态的高昂费用和进展缓慢,是由于普遍存在的组合效应、从体外模型推断到患者的困难、副作用以及药物耐受性/抗药性。目前的方法主要集中在通过使用CRISPR、基因治疗、蛋白工程或途径重构(mRNA和针对特定蛋白质的药物)来操纵细胞的“硬件”,以改变细胞机器和通信系统的个体组成部分。这种自下而上的方法通过微观管理对个体基因产物的改变存在一个关键限制,即逆问题的困难[2]:为了实现期望的系统级效应,需要调整哪些基因或分子?这个问题限制了基因编辑方法的应用范围,超越了单基因、线性疾病的低挂果实,因为在高度涌现、上下文背景敏感的复杂系统(如活体)中,没有通用的程序可以知道必须改变哪些微观层面的相互作用规则来实现特定的结果。
当前的分子水平方法类似于20世纪40年代和50年代计算机的物理重连方式进行编程。在信息技术革命中,计算机科学实现了令人瞩目的飞跃,通过专门的接口提供刺激来控制功能,以利用内置的高级模块化信息处理能力。多样智能和基础认知领域的最新进展表明,生物体在各种组织层次上都具备这种模块化信息处理能力[3-5]。生物医学能否从对分子硬件的摸索中实现类似的飞跃,发展出利用生命的这种生理“软件”的工具呢?对这种实时决策“软件”的几个显著特征(模块化、自上而下的控制和多尺度能力)的新认识,使生物医学工作者能够利用细胞和分子网络的集体智能[6,7]。
|
框1. 多尺度能力架构:嵌套的集体智能及对再生医学的影响
身体的一个显著特点是,问题解决能力在各种组织层次上都存在,并跨越不同的问题空间发挥作用。分子智能表现在执行趋化性的单个分子[149],以及形成由先前刺激模式驱动的记忆的基因调控网络和途径[7,24–26,150]。细胞骨架结构可以合理地编码记忆[151],细胞外基质(ECM)通常被细胞用作协调活动的信息素刮板,就像更熟悉的集体智能(如蚂蚁群体)使用环境一样[152]。因此,在生物医学背景下,细胞外基质细胞可以成为一个吸引人的介质,用于操纵细胞的感知和随后的行为。细胞智能表现出不同程度的能力,特别是在通过整合和优先处理来自微环境或自我生成的线索(如在体组织或工程迷宫中)导航复杂环境的能力,并根据感知历史做出决策[1]。组织通过改变基因表达来实现功能,尽管没有直接的进化适应力,以应对强大的毒素[22]。这是基于对先前共享某些共同特征的应激因子的响应进行问题解决和泛化的例子。在整个器官水平上,发育生物学充满了在发育和再生系统中通过解决问题实现复杂解剖终点的例子,尽管染色体组合、细胞大小和数量或大规模损伤发生了意外变化[6]。在生理、转录和解剖问题空间中,所有这些能力都实现了威廉·詹姆斯对智能的定义,即通过多种手段达到特定目标的能力。所有的能力水平都可以用于生物医学目的。在组织和器官水平上,简单的生理触发刺激可以诱导复杂的、自限制的器官发生,以实现再生[69]。在单细胞智能水平上,时间依赖的刺激协议可以利用药物的联想条件作用来打破药物耐受性(从而延长再生疗法的疗效)[26,138]。在这两个层次上,重设稳态设定点使有机体能够在没有持续干预的情况下保持健康状态[54,63]。除了向细胞传达目标和信号外,还有一个令人兴奋的机会,即通过使用人工智能工具来读取细胞网络的内部记忆和信念状态(类似于神经解码),并构建基于细胞生理/转录网络中强大的局部泛化和问题解决能力的计算模型[22],以确定与期望结果相关的基因和转录靶点(即利用细胞解决新问题的能力来帮助生物工程师解决限制CRISPR和类似技术应用的逆问题[2])。
|
二、身体智能:
解剖稳态、集体智能和功能需求驱动的修复
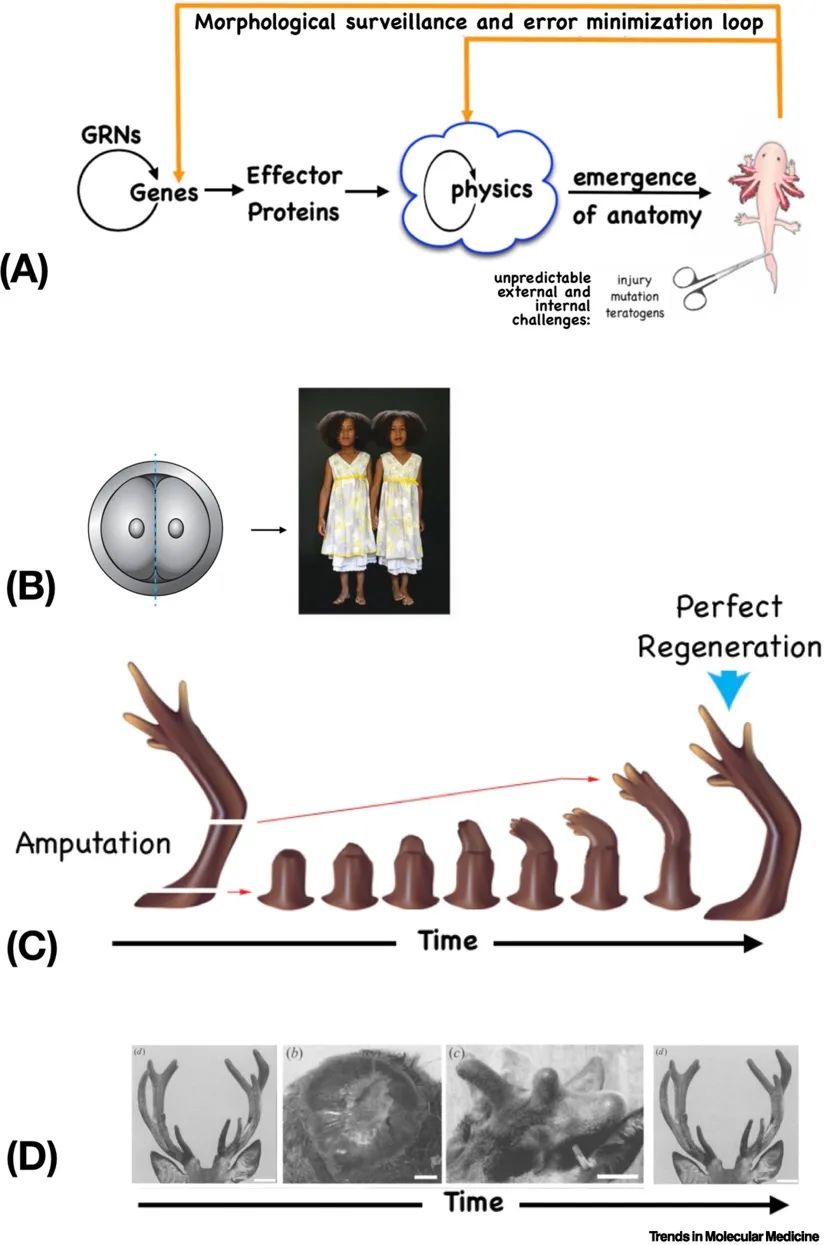
我们进化历史中最重要的一个方面,对医学有许多影响,就是我们由独立的单细胞生物进化而来。不仅是生活在我们体内的微生物,我们的实际细胞也具有感知、决策、记忆、预测和许多其他能力[1,8–12]。个体细胞在处理细胞层面的目标方面非常出色,例如在各种微环境扰动下保持适当的代谢和生理状态。关键是,当它们融入到多细胞生物中时,并没有失去这些能力。相反,进化将它们的最小愿望从单细胞的小设定点景观(增殖、代谢状态等)扩展到更大的组织和器官层面的目标:根据多细胞生物的功能需求构建特定的解剖结构[13,14]。
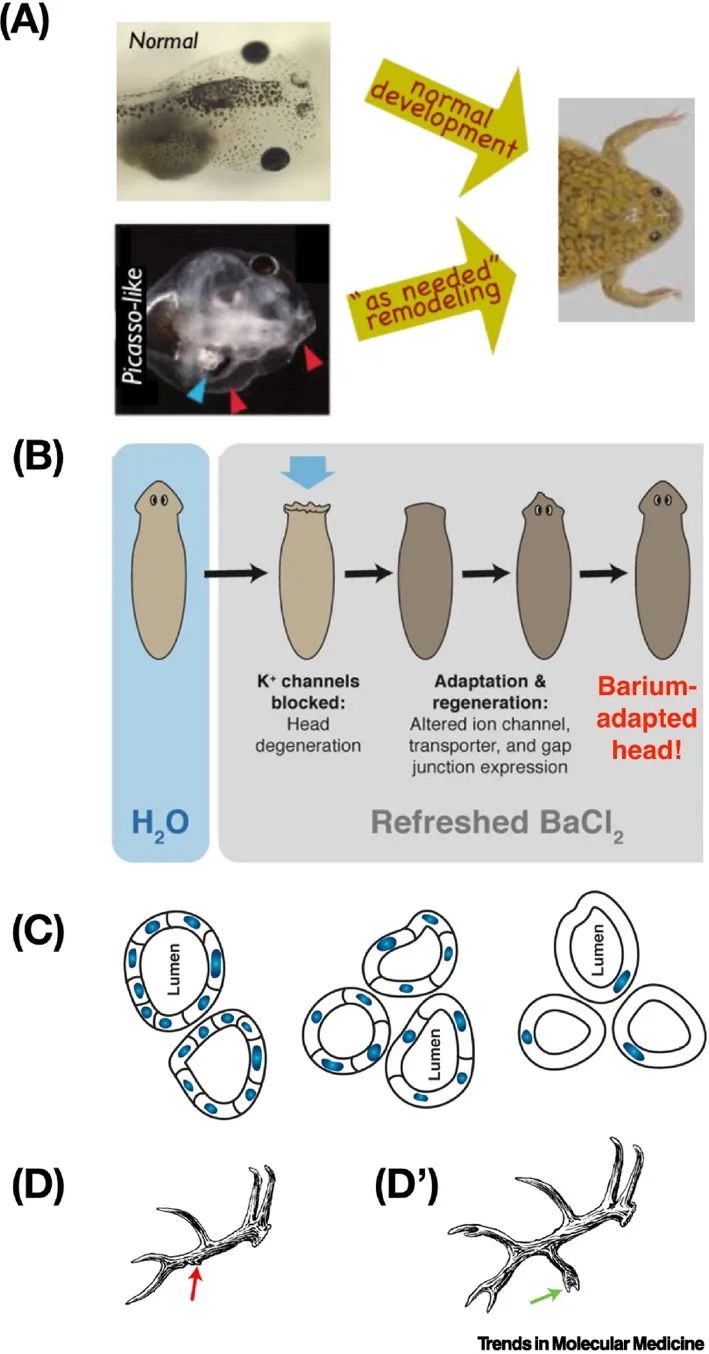
解剖稳态(anatomical homeostasis,通常称为“异稳态”)(图2A、B)的能力使人体能够准确发育和更新组织,以及一些生物体更令人印象深刻的能力,如再生整个附属肢和器官。例如,蝾螈可以再生肢体、眼睛、颌骨和卵巢(图2C)。甚至在成年哺乳动物中也有例子:鹿每年都能再生鹿角,重新生长大量的骨骼、血管、神经和皮肤,并具有形成新的形态记忆的能力(图2D)[15]。细胞能够迅速检测到与正常目标形态的偏差,并重建所需的结构,在正确的目标形态完成时停止。这也是早期胚胎分裂能够产生正常完整的单卵双胞胎而不是半个身体的原因(图2B)。研究追踪细胞集体在发育、再生和癌症抑制中如何部署群体智能表明,细胞群体和器官在变态发育过程中并不遵循固定的路径:相反,它们通过新颖的路径移动,直到达到目标解剖结构。例如,当蝌蚪的头颅器官混乱时,仍然可以形成正常的青蛙,因为这些器官会迁移,直到构建出正确的青蛙面孔(图3A)[16]。这种解决解剖和生理状态空间问题的潜在能力是任何干预措施中极其强大的组成部分的机械基础:让身体在事后愈合和适应。
关键是,细胞集体不仅可以在干预措施下达到相同的目标形态,还可以为新问题创造解决方案[6,17](图3)。从身体中分离出的上皮细胞可以重新启动它们的多细胞状态,并创建具有未经修改的青蛙或人类基因组但完全不同的身体结构、行为和能力的新型原生生物[18–21]。这种结构和功能命运的可塑性以及细胞集体的决策能力是管理复杂系统级结果的非传统且具有潜在变革性的目标。
这种自主性不仅仅是通过在解剖形态空间中解决问题来确定身体/器官的形状。在生理和转录空间中,关于对新压力的创造性解决方案的显著例子层出不穷,并且这些例子在模型系统中开始被理解。例如,当扁虫接受钡处理(一种非特异性的钾通道阻断剂)时,它们的头部迅速退化;然而,如果继续处于钡环境中,它们很快会重新生长出对钡不敏感的新头部(图3B)[22]。这是通过诱导一小部分基因来实现的,这些基因有了新用处,即在钾通道功能被废除的情况下保持一个可操作的头部。由于扁虫在自然界中从未遇到过钡,这不是一种进化解决方案的激活-这使得组织能够准确地识别出解决这种生理应激的正确基因更加引人注目。类似地,即使在实验中管道细胞的大小发生了巨大变化,蝾螈肾小管仍然会发育到指定的直径。当推向极端时,细胞形成管道的机制甚至会改变以保持目标形态(图3C)。在生理和转录空间中这种能力的全部范围尚不清楚,但它们很可能由所有智能行为的已知组成部分实现:各种形式的学习,这些学习现在已经在非神经组织中被发现[7,23,24],甚至在身体中普遍存在的亚细胞组分,如基因调控网络和信号通路[7,24–27]。这些能力现在开始在临床环境中得到利用。
(三)模式稳态和异稳态:组织发育、维持和再生的统一框架
发育、再生和抑制癌症可以被视为同一普遍、动态过程的不同方面:模式稳态。这是为了根据编码的设定点维持一组特定的功能解剖特征。当蝾螈的腿被截断后,它会再生,细胞持续增殖和迁移,直到形成一条正确的腿,然后停止。细胞集体通过持续的、主动的误差最小化来实现这种模式稳态,这在寻找组织抵抗癌症和衰老的组织原则方面已经被广泛讨论[28],并且在信息缺失导致已形成结构的紊乱的例子中(例如,切断舌神经导致舌乳头的紊乱[29])。框2将详细阐述这个想法。
|
框2. 发育和再生作为上下背景文敏感的问题解决过程
胚胎发育不仅仅是机械细胞水平规则的不可避免的、紧密联系的工作,而是一系列“修复”每个胚胎阶段,朝着更正确的下一个阶段发展的再生事件。这个过程由快速变化的生物电预模式引导,这些预模式编码了目标形态,根据这个目标形态估计和最小化误差[14]。这种不断变化的目标状态是异稳态的一个例子[157–162],其中二阶机制逐渐改变设定点本身,从而在成熟和变态过程中实现有向变化。类似地,计划虫会按比例缩小和增长,并不断重塑身体,使其与可用细胞数量完美匹配[163];脊椎动物胚胎也是如此,在身体大小增加时扩展肢体和器官[164]。
活体在决策方面具有极高的能力,可以在外部环境和内部组分的巨大变异性下优化功能。计划虫是完美的再生者,同时也是不老和抗癌的,尽管其基因组和混合倍体性非常混乱[165]。蝾螈即使细胞因多倍体而变得巨大,也能产生正确大小的身体结构[156]。整个附属物可以自我纠正:移植到侧腹部的蝾螈尾巴会慢慢重塑为肢体——这是一种更适合大尺度解剖环境的结构[166]。因此,大尺度的解剖和生理规范驱动着潜在的分子生物事件。
这些现象的核心是生命的上下文敏感的问题解决能力,超越了基因确定的蛋白质硬件。在纤毛虫皮质遗传中发现了非遗传的模式记忆[167];基因组之外的信息(以及其许多染色质修饰)在细胞骨架结构和其他基质介导下,延伸到鹿鹿角解剖学的营养记忆[2](图3D,D′)和计划虫再生[169]。在突变了semaporin(神经引导信号)蛋白的小鼠中,错位的背侧外侧膝状核(dLGN)轴突使用替代路径找到通向视觉皮层的路线[154],类似于蝌蚪变态中混乱的颅颜器官[170]。学者Slijper的山羊提供了一个经典的例子[171],在这个例子中,一个缺乏前肢的哺乳动物在发育(而不是进化)的时间尺度上获得了许多为二足动物运动所需的解剖调整。这些动态的大尺度行为由一个执行“有机体需求”的计算的生理层实现[172,173]。我们将在发育生物电部分回到这个生理层,但首先提供一个关于哺乳动物肝脏细胞和组织级别对功能变化的响应能力的临床例子。
|
这种观点认为,细胞集体积极导航解剖、生理和转录空间以实现目标,这在体内得到了几个预测的验证。其中之一是,即使是重大的分子干扰,例如扰乱左右体对称性设置机制的治疗,也会随着时间的推移自我纠正[30]。这种纠错能力既是潜在的故障点,当受到压力破坏时会导致复杂疾病,因此非常适合调节的目标。
三、“肝稳态”:
细胞智能及其在肝病临床应用中的例证
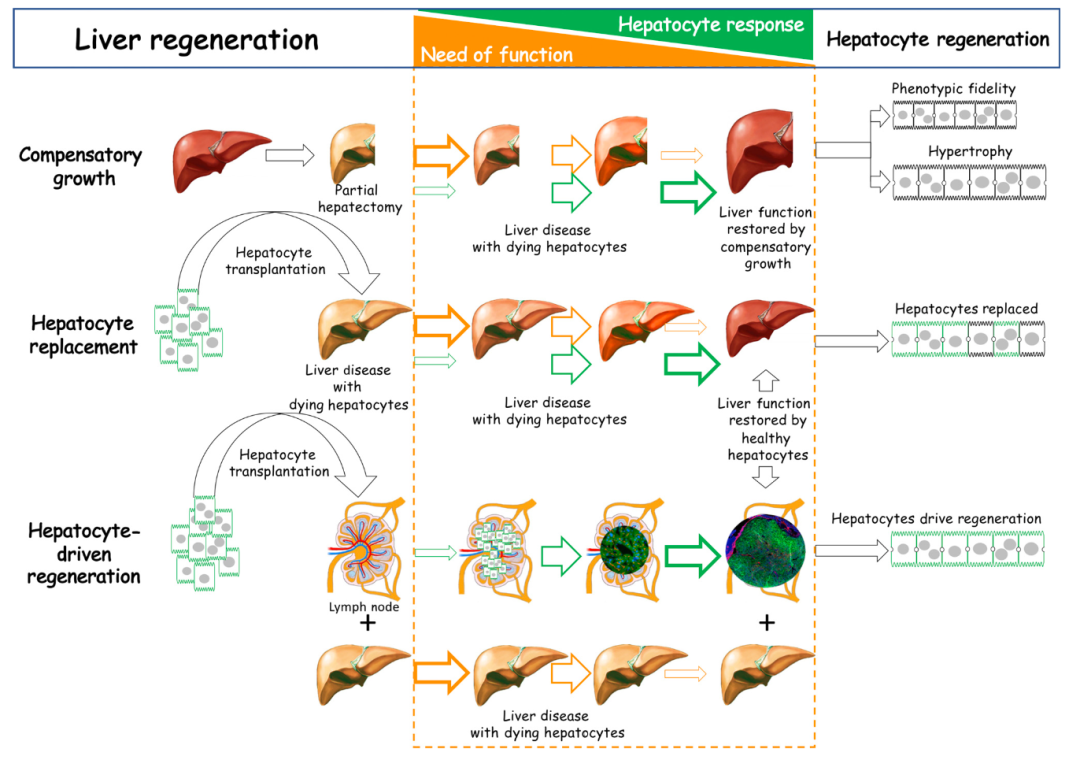
慢性肝病的全球负担不断增加(在美国,它是45至55岁人群死亡的第四大原因),而治疗选择有限[31],这使得它成为再生医学创新的重要目标。肝脏具有非凡的再生能力,这是从鱼类到哺乳动物的脊椎动物的关键特征[32]。除了数百种生存所需的功能,包括解毒、消化、过滤、新陈代谢和蛋白质合成,肝脏还充当免疫系统的守护者[32]。作为人体最大的器官,肝脏的大小受到严格调控,以维持正常的葡萄糖和氨水平等生理功能。肝脏是唯一一个利用再生机制来确保肝脏与体重之比保持恒定的实体器官。这种稳态特性(图4)——“肝稳态”[33]及其再生潜力——是肝脏独特的特性,其特性表明它是具有高度临床相关性的细胞智能的例证。
肝脏再生是复杂的,因此多种再生机制的进化确保了肝脏功能的稳定性[35]。自19世纪以来,通过部分肝切除手术移除肝脏组织已经得到广泛研究。与蝾螈或蝾螈能够再生整个附属肢和器官不同,剩余的肝脏会经历快速的代偿性增生,以恢复原始的肝脏质量,但不会恢复多叶状的肝脏解剖结构。此外,它还显示出表型的保真性:肝细胞生成肝细胞,内皮细胞生成内皮细胞,等等。有趣的是,代偿性生长是通过增殖和肝细胞大小的增加的组合来完成的[36]。在70%肝切除术后,60%的肝细胞会进行分裂,但在30%部分肝切除术后,没有细胞进行分裂:仅仅增加肝细胞的大小就可以弥补细胞的损失,并且是增加肝脏功能的更快更有效的方式[36]。
肝细胞移植长期以来一直被提出作为正位肝移植(OLT)的潜在替代方法,因为移植的肝细胞可以恢复缺失的肝脏功能。然而,这种方法对于100多名肝病患者只提供了部分矫正[37]。为了取得成功,移植的肝细胞必须替代疾病肝细胞以弥补功能损失,而这种损失因疾病而异。在尿激酶纤溶酶原激活剂(uPA)转基因小鼠中的早期结果(其中细胞内uPA的激活导致肝细胞损伤和死亡)显示了肝细胞替代的前景,但同源重组和毒性转基因的删除降低了移植的成功率[38],这是有效肝细胞替代的关键。在遗传性酪氨酸代谢障碍(HT 1)的小鼠和猪模型中取得了更好的结果,HT 1是一种由于酪氨酸代谢途径中FAH基因的突变或缺失,导致的婴儿期逐渐发展的肝脏损伤的人类遗传性疾病[39]。在这些模型中,健康肝细胞的移植在6周(小鼠)至3个月(猪)内替代了肝脏中超过90%的疾病肝细胞,并拯救了动物免于致命的肝脏疾病。因此,供体来源的肝细胞可以在肝脏微环境中成功与疾病肝细胞竞争。
图4 以肝细胞为中心的肝脏再生和功能需求视角。在部分肝切除后,肝细胞通过增殖、表型保真和肥大的方式,响应功能需求,恢复肝脏质量。在肝细胞移植到患有肝细胞疾病的肝脏时,健康的肝细胞取代不健康的肝细胞,以恢复肝脏功能。
细胞移植已被提出作为正位肝移植的治疗替代方案,或者作为等待正位肝移植的患者的过渡措施[37]。然而,移植的人类肝细胞通常通过脾动脉或门静脉注射,对于绝大多数患者来说,肝硬化/纤维化及门静脉高压使得这些途径具有危险性或不可行性,可选择的选项非常有限。辅助肝的概念被提出作为一种替代策略。通常情况下,健康的肝脏移植物被放置在异位或正位,使得原有的疾病肝脏全部或部分保留。这种方法不仅有潜力为移植患者提供过渡,还可以通过接受原有肝脏的自发再生潜力来避免进行正位肝移植。尽管早期试验中存在问题,但在急性肝衰竭[40]、代谢紊乱[41–44]甚至肝硬化[45,46]的病例中,最近更有利的结果令人鼓舞。
(一)通过异位肝脏利用肝细胞的计算能力进行功能补充
在诱导肝衰竭的酪氨酸代谢障碍小鼠中,将肝细胞移植到非肝脏的异位位置后,发现了制造新的辅助肝的概念:肝细胞在淋巴结中植入后再生出功能性的辅助肝。分析显示,肝细胞是唯一植入淋巴结的供体肝细胞,并随后分裂并吸引宿主的其他细胞。这表明肝细胞充当了乐团指挥,通过引导包括宿主细胞在内的细胞反应来生成功能性的辅助肝。辅助肝的生长似乎也是由功能需求驱动的:一旦达到足够的肝功能质量,肝细胞的增殖就会停止[47,48]。然而,如果对原有肝脏进行部分肝切除,淋巴结中的肝细胞会再次开始增殖,直到辅助肝达到正常肝脏质量和功能。值得注意的是,在没有严重肝脏疾病的情况下,移植到淋巴结的肝细胞不会生成肝组织[48],支持了功能需求的概念。将肝细胞直接注射到酪氨酸代谢障碍小鼠的单个淋巴结中,可以生成约70%正常原有肝脏质量,足以拯救这种异位肝脏的表型。异位肝脏无法达到100%正常原有肝脏质量,因为原有肝脏仍然存在并具有一定的肝功能[47,48]。
最近的转录组分析比较了辅助肝和原有肝的差异,发现辅助肝中选择性地表达了控制肝功能的基因,暗示着两个肝脏之间存在调节的功能整合[49]。正如预期的那样,在原有的酪氨酸代谢障碍肝中,FAH基因和其他四个编码酪氨酸降解途径酶的基因,其表达被显著抑制,而辅助肝的表达水平与野生型对照组中的肝脏相似。类似的结果也得到了调节凝血系统、尿素循环和白蛋白合成的基因的研究。这些观察结果再次强调了肝细胞功能的复杂调节,以在不同条件下维持肝功能的稳态。
大型动物研究提供了将肝细胞移植到淋巴结作为治疗肝衰竭的临床前概念证明。在酪氨酸代谢障碍的FAH-/-猪中,自体FAH-/-肝细胞被分离出来,经过离体转导使用携带人类FAH cDNA的慢病毒载体,然后移植到肠系膜淋巴结[50]。肝细胞在淋巴结中的植入在移植后的6小时内观察到,并持续超过8个月;在移植后的97天,已经检测到足够的肝脏质量以缓解急性肝衰竭。在解剖检查中,淋巴结中存在具有肝叶的辅助肝,并且已经确定了血管化,但也检测到了FAH基因转导的肝细胞对原有肝脏的再充实。追踪注射的转基因细胞显示,肝细胞的初始扩增发生在淋巴结中,生成了健康肝细胞的来源,随后再充实了患病的肝脏[50]。进一步支持功能需求的概念,一旦原有肝脏被健康肝细胞恢复,辅助肝的大小会减小并最终消失,这与之前在酪氨酸代谢障碍小鼠中的观察结果一致[47]。在外杂种猪模型中,手术诱导的亚急性肝衰竭也得到了类似的结果[51],这表明对于异位肝细胞的功能需求调节并不特定于酪氨酸代谢障碍。
利用淋巴结作为移植的场所,肝细胞驱动的功能需求和辅助肝的生成的蓝图已经应用于其他器官。胸腺、胰岛、肾脏和其他几种组织显示出了潜力。解决内部或外部引发的压力和损伤的关键是利用细胞的适应能力。医生对于pH值或温度等标量属性的稳态非常了解,但前文总结的工作和类似的证据表明,机体对于适应性稳态的利用超越了这一点,并为治疗干预提供了有希望的目标。迅速积累的证据表明,机体的所有组织层次都包括参与实时控制和原型认知信息处理的控制系统,例如感知控制回路和最小化意外[52,53]。这些系统为生物医学管理提供了一条新的路径:以期望的结果为目标,针对稳态设定点[54]和细胞感知机制[1,8,9,55,56]进行干预,让稳态系统发挥其最佳功能——以灵活和上下文背景相关的方式实现其目标。
四、利用身体智慧调节再生:
神经生物学和行为科学指引方向
细胞群体中的集体智慧协调着细胞行为,实现了复杂的健康器官,尽管细胞不断更新、受伤和面临新的压力。是什么系统使这成为可能?我们如何理解和利用这种集体智慧来进行生物医学研究?在生命的多尺度能力架构中,每个层次(从亚细胞到器官系统再到社会群体)解决了自己特定的问题,细胞和细胞集体以智能的方式在转录和解剖空间中导航,就像动物以不同程度的智能在三维空间中导航一样[57]。我们如何利用这一洞见进行生物医学控制?行为科学和神经科学这些成熟的领域已经通过设计影响内部决策的输入和刺激,提供了理解、预测和操纵极其复杂系统的途径。这些领域在多尺度方法上有着悠久的历史,涵盖了从突触通路机制到精神病学等多个层次。这些可以作为生物医学的模板,使其在当前对越来越低层次目标(基因、蛋白质、通路)的关注之外扩展。
神经科学通过关注处理信息以实现适应性目标的机制和算法来解决问题解决和智能。相比之下,行为科学则从另一个尺度入手。在我们对大脑细节一无所知的几千年前,人类就能够通过训练来控制动物。这说明可以在机制细节清晰之前,就实现强大的控制,估计系统的智能程度,结合研究作为奖励和惩罚的刺激和状态。行为科学不仅提供了一条不需要完全了解底层细节的系统级结果的路径,而且显示了这种方法的效率:即使通过管理所有神经信号来自下而上地控制动物的行为是可能的,但通过训练从上而下地实现这一点更加高效(并提供更灵活的结果)——利用系统的本能感知记忆能力。
同样地,利用身体重新调整其内部状态以实现期望的结果的能力,将使我们能够解决困扰医学的复杂性问题——一切都与其他事物有一定的联系[58-60],因此很难得出简单、可靠、线性的干预措施。幸运的是,细胞和组织提供了一个内置系统,可以重新调整它们自己的复杂分子状态,以实现系统级目标:学习。学习通过管理无数的分子细节来实现所需的器官或整体身体规模的反应,完成了困难的任务。从短期来看,目前的生物医学方法无法实现这种类型的控制,这种控制是由感觉或受体层的高级体验引发的。然而,这为生物医学干预提供了一个强大的接口:有人认为,未来处理复杂疾病的许多方法将采取训练和操纵通路输入的形式,而不是重构它们[61-63]。
五、发育生物电:
与局部高级解剖控制系统的操作接口
直到最近,学习被认为是大脑和神经系统的独特能力——为什么我们会期望同样的技巧在身体其他部位起作用呢?大脑的卓越信息处理能力从何而来?大脑的所有分子机制——包括离子通道、电突触和神经递质,以及许多指导体内生物电网络功能的算法——都是古老的[64,65],并且在整个身体中都被使用(图5A-D)。进化在微生物生物膜出现时发现了生物电网络的计算能力[66,67],并一直在利用这一能力。细胞表面的离子通道设置细胞的静息电位(Vmem),并且可以通过称为间隙连接的电突触与邻近细胞进行通信,从而创建区域生物电网络(图5B)[68]。几乎身体的每个组织,不仅仅是神经元,都形成这样的生物电网络,现在学界已经认识到这些网络的输出控制细胞的增殖、分化、迁移和凋亡等行为[69]。就像神经网络通过控制肌肉状态来处理信息,使身体能够在三维空间中实现行为目标一样(图5C),这些非神经生物电网络通过控制细胞增殖、迁移和分化(发育生物电)来实现解剖形态空间中的形态目标(图5D)[57]。控制神经元和体细胞电生理状态的机制的相似性使得神经科学的技术可以应用于非神经生物电网络(图5E)。很可能,使大脑能够通过高效的高级相互作用进行控制的特性并不是独特的——而是进化根据需要将通用细胞能力加速和修改为不同问题空间的版本[14,70]。
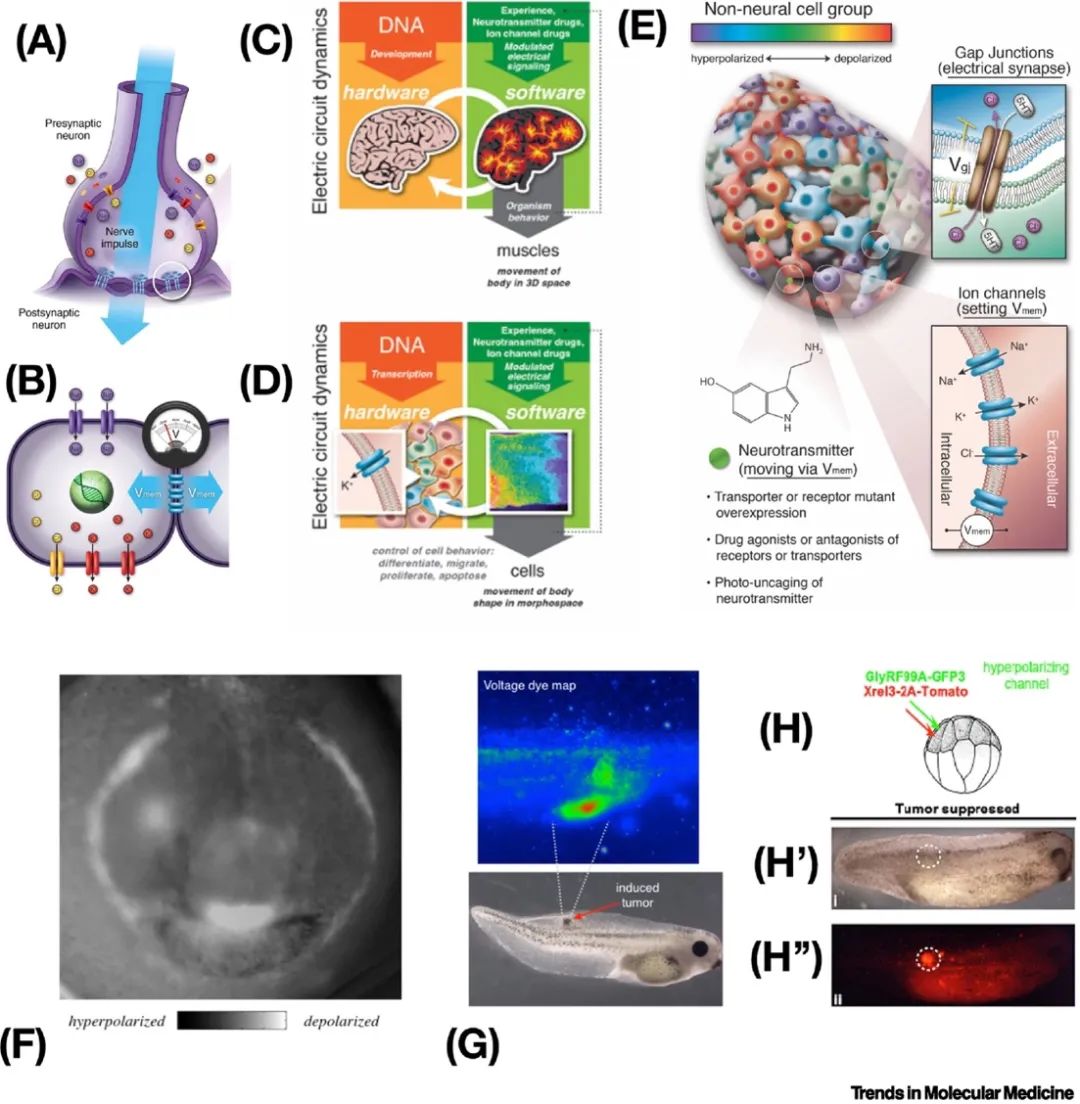 指导后续基因表达和解剖的内源性生物电预模式,例如青蛙胚胎中的“电脸”(图5F)[71],对于正常发育是必需的,并编码了细胞将构建的目标形态。远程生物电决策已被认为参与了附肢的形成、特定器官的诱导、大小控制和主要体轴的对齐[72-74]。因此,操纵生物电界面在动物模型中已被证明对刺激伤口愈合[75-77]、附肢再生[78]和修复先天缺陷有效。现有的计算方法可以预测使用经过人类批准的离子通道药物(电药物[79,80])的有效治疗方案,这些药物可以在动物模型中触发对大脑、心脏和肠道的复杂畸形进行修复[81-83]。令人惊讶的是,这种修复不仅适用于由化学致畸原引起的畸形,还适用于由关键调控基因Notch的突变等遗传紊乱引起的畸形[81,83]。这些结果表明,即使是一些具有多效性和复杂效应的遗传硬件缺陷,也可以通过相对简单的触发刺激在“软件”层面上进行修复。在癌症研究中也观察到了类似的遗传默认状态的改变[84-87]。尽管存在强大的致癌突变,如KRAS变异体(图5G,H),但离子通道调节可以驱动细胞朝着正常表型发展,从而在体外和体内实现肿瘤的正常化或预防。
为什么生物电状态能够控制复杂的结构和功能?为什么神经科学的主要工具(如光遗传学、主动推理框架和精神活性化合物)在大脑之外的地方以类似的方式起作用?最近的研究表明,生物电是一种“认知胶水”,将个体细胞与整个身体的共同器官目标联系在一起[13,88];生物电网络不仅存储解剖稳态的设定点,还实现了组织识别大规模错误并追求正常化所需的分布式通信和计算。已经证明,模式记忆(存储形态发生学将尝试构建的稳态设定点)以稳定的生物电模式形式存储,可以被可视化、解释和重写[72]。因此,例如,可以编辑扁虫的存储目标形态,使其读取“两个头”而不是默认的“一个头”,并产生一系列永久再生的双头扁虫,而不改变其基因组。
至关重要的是,与大脑类似,体细胞的电生理状态为进行大规模决策的计算介质提供了特权访问。这一新兴领域的进展揭示了结果不仅仅是前馈性的紧急情况,而实际上是一个具有明确编码、可追踪目标状态的控制过程,为解决限制自下而上干预的复杂性、非线性和退化性问题提供了一种方法[2]。因此,生物电为基因组指定的微观分子硬件和大规模解剖结果之间提供了生理控制层,是高级生物医学干预的理想目标。
指导后续基因表达和解剖的内源性生物电预模式,例如青蛙胚胎中的“电脸”(图5F)[71],对于正常发育是必需的,并编码了细胞将构建的目标形态。远程生物电决策已被认为参与了附肢的形成、特定器官的诱导、大小控制和主要体轴的对齐[72-74]。因此,操纵生物电界面在动物模型中已被证明对刺激伤口愈合[75-77]、附肢再生[78]和修复先天缺陷有效。现有的计算方法可以预测使用经过人类批准的离子通道药物(电药物[79,80])的有效治疗方案,这些药物可以在动物模型中触发对大脑、心脏和肠道的复杂畸形进行修复[81-83]。令人惊讶的是,这种修复不仅适用于由化学致畸原引起的畸形,还适用于由关键调控基因Notch的突变等遗传紊乱引起的畸形[81,83]。这些结果表明,即使是一些具有多效性和复杂效应的遗传硬件缺陷,也可以通过相对简单的触发刺激在“软件”层面上进行修复。在癌症研究中也观察到了类似的遗传默认状态的改变[84-87]。尽管存在强大的致癌突变,如KRAS变异体(图5G,H),但离子通道调节可以驱动细胞朝着正常表型发展,从而在体外和体内实现肿瘤的正常化或预防。
为什么生物电状态能够控制复杂的结构和功能?为什么神经科学的主要工具(如光遗传学、主动推理框架和精神活性化合物)在大脑之外的地方以类似的方式起作用?最近的研究表明,生物电是一种“认知胶水”,将个体细胞与整个身体的共同器官目标联系在一起[13,88];生物电网络不仅存储解剖稳态的设定点,还实现了组织识别大规模错误并追求正常化所需的分布式通信和计算。已经证明,模式记忆(存储形态发生学将尝试构建的稳态设定点)以稳定的生物电模式形式存储,可以被可视化、解释和重写[72]。因此,例如,可以编辑扁虫的存储目标形态,使其读取“两个头”而不是默认的“一个头”,并产生一系列永久再生的双头扁虫,而不改变其基因组。
至关重要的是,与大脑类似,体细胞的电生理状态为进行大规模决策的计算介质提供了特权访问。这一新兴领域的进展揭示了结果不仅仅是前馈性的紧急情况,而实际上是一个具有明确编码、可追踪目标状态的控制过程,为解决限制自下而上干预的复杂性、非线性和退化性问题提供了一种方法[2]。因此,生物电为基因组指定的微观分子硬件和大规模解剖结果之间提供了生理控制层,是高级生物医学干预的理想目标。
所提出的研究计划旨在弥合分子生物学家和合成生物学家之间的鸿沟,前者将生物体视为机器,而后者则持有有机主义、整体论的系统生物学、生态学和以人为中心的医学观点。活体是“机器”,并不是以天真的机械方式来理解,这样会剥夺它们明显的智能和主动性,而是指“机器”是一种可以被理性理解、修复、改进和创造的东西,只要找到与之交互的适当层次的主动性、有效的接口和对其内部有意义的刺激[14]。
将生物体视为多功能、多尺度的问题解决主体,可以利用行为和认知科学的工具来重新定义激进再生医学中的可能性。它为再生医学工作者提供了一组新的生物医学操纵目标,包括模块化多尺度控制、适应性调节回路、(非传统的)主体的内部信息状态,以及高度有效的问题解决机制。目前已有的可以通过短暂信号修复的结构缺陷,只是即将到来的再生医学路线图的冰山一角。在这个路线图中,干预将利用生物体的所有固有子程序和能力。从生理性疾病(如被视为一种学习障碍的糖尿病[89])到先天缺陷[81]、癌症[85]和再生能力[90],我们认为通过触发、重定向、修改和最终增强机体的自我监测和自我修复能力,可以解决非常多样的疾病条件。
具有真正集中式大脑、双侧对称和学习能力的扁虫为我们展示了可能性的一瞥。它们不仅具有很强的再生能力,并且在头部再生过程中保持记忆,而且还揭示了当前范式中的一个关键差距。根据当前理论,癌症和衰老是由于遗传错误的积累,扁虫的混合体细胞体应该充满肿瘤和衰老。然而,它们对癌症高度抵抗的且稳定的[91,92]。我们提出[93],这是由于进化过程中对生理算法的关注,这些算法在分子水平上存在极端噪声的压力下,需要产生高度适应性和功能性的身体。与当前理论认为未分化(即再生能力)的细胞对长寿动物来说是癌症风险不同,我们认为解剖可塑性控制机制和算法使得长期健康寿命和从损伤和压力中恢复成为可能。扁虫适应新的内部和外部挑战的能力是极端的,但所有生物都在某种程度上具备这种能力-即在不可预测的情况下努力寻求一致的解决方案的能力。理解和利用这种能力是再生医学的前沿。
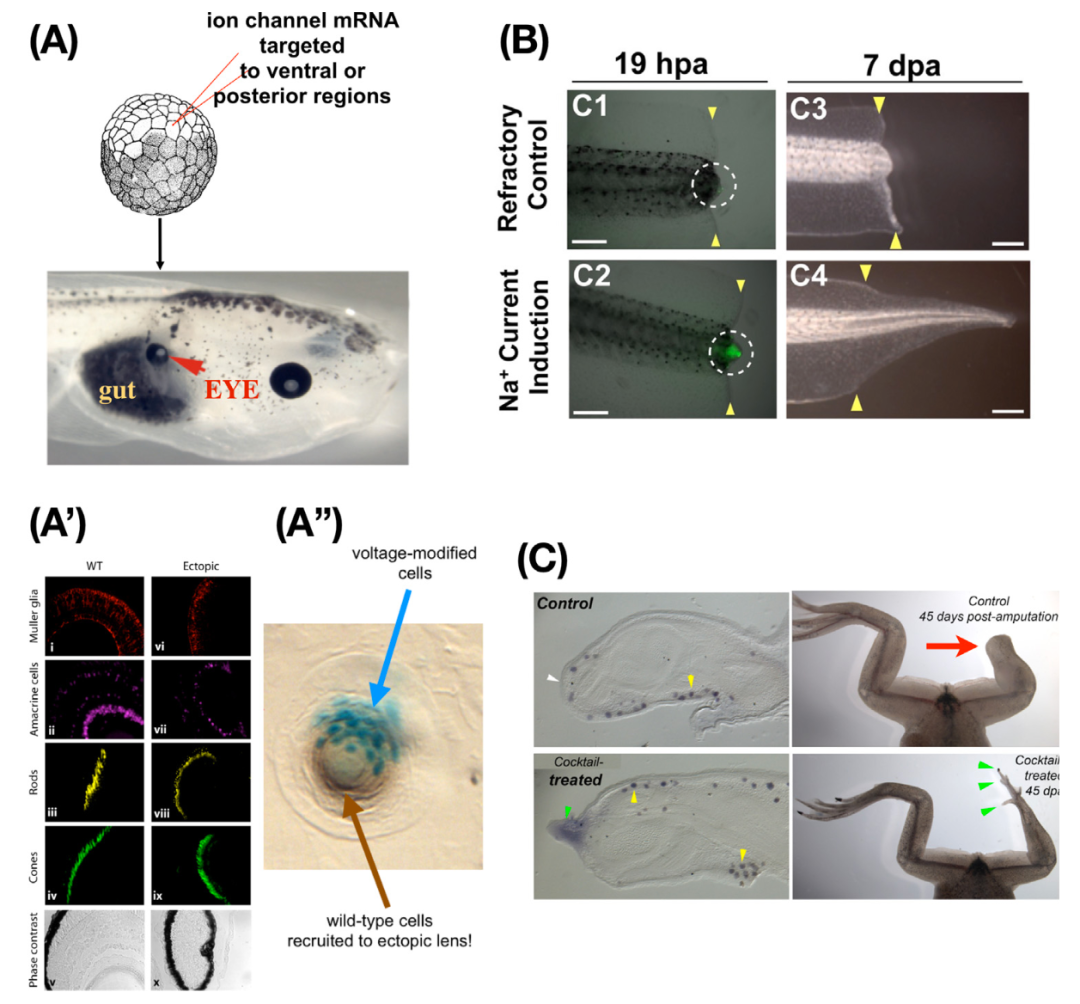
在临床前模型中,已经看到了未来医学的一瞥,其中利用生理信号重新编程肿瘤(图5H)[94-96]和诱导附属物再生。图6展示了通过精确操纵电生理状态来启动复杂的细胞行为子程序的能力的示例,包括在蛙卵中诱导异位眼睛(图6A)和蝌蚪尾巴和青蛙肢体的再生(图6B,C)。尤其引人注目的是,在前者中,即使在尾巴中诱导眼睛,它们也是功能性的,它们连接到脊髓而不是大脑[97,98]。此外,当处理的细胞数量较少时,仍然会产生一个功能性的眼睛,并且生成的晶状体包括处理过的细胞和被招募来帮助完成任务的未经操纵的邻近细胞。这种通过利用其他细胞来实现大小控制的能力是组织的固有能力,无需进行工程或微观管理,就可以在生物医学中加以利用。
利用集体细胞智能为生物医学开发复杂结果的潜力,而无需微观管理棘手的复杂性,是其独特而强大的优势之一。具体而言,通过识别目标状态的编码方式以及其存储和解释的方式,可以通过下游机制的简单触发(与身体的比例和方向、正确的内部结构和自限制生长)诱导整个器官(如眼睛和肢体)的形成。另一个新颖且有前景的例子是附属物(尾巴和肢体)再生,其中非常短暂的暴露[90,99]足以改变下游的决策过程。这种方法的一个关键优势是最小化意外后果(例如,当前分子水平方法的副作用)。通过将控制复杂性转嫁给生物系统本身,并利用固有的高度协调的程序,而不是试图微观管理所有必要的细节,可以避免许多意想不到的局部调整的后果。朝着临床治疗的关键下一步是全面了解体内信息处理能力的层次结构-了解细胞集体测量什么,它们如何在物理模态之间分配注意力,以及可以利用的其他能力。
(二) 互操作性:保持细胞问题解决能力的一个重要结果
理解这一现象的影响远不止于修复标准器官的形态和功能。细胞和组织解决新问题的能力是生命的互操作性的基础——它使得各种混合、杂交、生物工程、合成和机器人生物技术成为可能[17]。将藻类插入蝌蚪的大脑使其能够通过光合作用模式运作[100,101],智能植入物[102,103]能够轻松融入组织,闭环控制系统使新型生物能够在虚拟世界中运作[104,105],而当蝌蚪的尾巴上出现眼睛时,它们在视觉训练中表现出色[97,98]。这些都不需要传统意义上的进化适应,即通过基因介导和选择来实现:生物通信接口和问题解决能力足够通用,以至于它们可以立即在不同界域(神经元和细菌、大脑和藻类等)和进化的自然形态以及设计的无机结构之间发挥作用[106,107]。未来的生物工程和生物医学无疑将在伦理考虑的背景下,利用这一强大的普适设计原则,这包括改善健康的必要性以及合成生物工程的影响等更广泛的问题[17,108,109]。
几种理解生命体复杂性的框架[14]表明,存在一种计算能力的范围,其中不仅包括机械系统,还包括具有稳态、适应态、学习和更高级能力的系统。这形成了一种可说服性的连续性——一个光谱,在这个光谱上可以使用几类概念和实证工具来诱导所需的变化(图7A、B)。我们不能简单地猜测生物体的组成部分在这个连续性谱上:我们必须进行实验,并准备好对于关于细胞新能力的发现,以及发现细胞抵抗操纵方式的意外的行为复杂性。一个引人注目的例子是信号通路和基因调控网络。尽管这些确定性的简单系统似乎是机械钟表的典型案例,只能通过重连来进行修改,但它们已被证明具有六种学习类型[25,26]。此外,这些网络可以像有智慧的动物一样高效地受控——通过使用像巴甫洛夫条件反射等训练要求[24-26]。除了之前描述的稳态目标重设策略,生物组织还提供了哪些有效的预测和控制的机会?
身心医学(Somatic psychiatry)描述了一旦我们将身体组织视为具有多样化问题空间导航能力的集体主体,就可以将行为和认知科学的技术视为干预手段。最近在高级患者信念和药理途径交叉领域的进展[110–113],开始显示了对身体生物化学的自上而下控制的重要性。生物控制的多尺度性不仅限于安慰剂效应。即使是早上自愿起床也是一个过程,其中执行功能(高级认知状态)最终控制肌肉细胞的膜电位(分子水平的生物物理特性)。因此,身心医学(框 3)[114–116]并不是一种罕见现象——相反,它是适应性行为中不可或缺的日常事件,并为健康和疾病的变革性方法提供了深刻的机会。行为神经科学提供了一个框架,用于理解高级心理状态和物理状态相互作用的多尺度现象:它具有适用于从突触蛋白到社会精神病学的组织层次的处理方法,以及跨层次框架的先例[117,118]。
|
当前生物医学和生物工程方法的重点主要是自下而上——通过控制最低级组件(蛋白质、DNA序列等)来实现特定功能。然而,生物学使用了一个集成的、多尺度的能力架构,其中更高层次的组织决策关于我们想要控制的系统级结果——大规模形状和复杂生理状态。最典型的例子是在神经系统中,认知状态(目标、信念、希望、意图等)必须与身体的功能性相连接。关于生物反馈、身心医学(例如,冥想实践或暴露于音乐后大脑基因表达的变化)、心身免疫学和安慰剂/无效剂效应的最新和经典研究已经清楚地表明,生理和基因状态可以由高级节点控制。需要注意的是,身心控制并不是一些特殊情况下的异常情况,比如催眠状态。每当一个人早上起床开始一天的任务时,使这一切成为可能的是一种多尺度传导机制,将执行级元认知意图转化为肌肉细胞静息电位的去极化。因此,我们的具身心智已经具备了控制复杂分子事件并将其引导到适应性行为的能力,而不需要每个层次都了解其下方和上方的细节。Fabrizio Benedetti等先驱者的工作[110–113]表明,药物暴露和对药物的期望激活了相同的机制,这证明了我们进化的架构的一个关键方面,可以在治疗中加以利用。认知状态实施复杂的下游变化的能力不是大脑的独特特征,相反,智能和分布式控制融入了所有体细胞和组织中,并且是潜在的治疗目标。将复杂目标(例如,生长一个适当大小和形状的器官)与分子实施机制相连接的局部生物电界面开启了一个变革性的可能性,即人工智能可以作为强大的GPS,引导对生物组织的控制,以导航转录、生理和解剖景观。新的进展,如大型语言模型,提供了一种可能性,即它们可以成为我们思维(以及诱导健康的目标)与身体原始智能之间的翻译者,通过帮助推导刺激、训练协议和经验作为塑造生理和解剖子系统行为的治疗方法,以增加健康寿命。
|
具体而言,我们建议应该越来越多地研究组织、器官和分子途径的多种学习和原型认知能力。很可能基础认知和单细胞训练的文献[119,120]可以发展成为训练细胞达到所需基因表达水平、生理状态或解剖结果的方案。已经有努力将糖尿病视为学习障碍[89]和将心脏记忆建模为真正的记忆[121]。此外,现有的可通过自上而下控制的认知层次之间的界面可能被利用来控制生理和生化的身体状态[122–124]。在某种程度上,这已经开始了,通过生物反馈[125]、安慰剂/无效剂研究[126,127]和催眠皮肤科学[128]的努力。在所有这些情况下,高级认知信息被转化为细胞和分子水平的变化。
自从巴甫洛夫以来,人们就已经知道生理回路可以通过经验或在机体中传播或扩展的持续生理状态进行训练[129]。一个例子是,健康的对侧腿迅速显示出与受伤腿相同的生物电状态,位于相同的位置[130],揭示了相对特定的状态信息在整个身体中的传播。再生医学方法可以通过使用临时的生理刺激来控制细胞行为并诱导再生修复,从而与物理损伤和应激因素设置持久的生理状态相类似。正如前面所述,动物和细胞模型的研究已经证明,在胚胎发育和再生中协调细胞集体行为的生物电控制系统,是逆转肿瘤的可操作接口[84,85]。由于体内的体细胞生物电系统与大脑中的系统之间存在相似之处,因此有可能通过针对生物电系统,来提供比重写稳态设定点更强大的控制身体过程的方法。这将使得行为科学的技术(针对信念、记忆和预期的自我和世界模型)能够控制复杂的系统级结果,这些结果对现有方法具有抵抗力。用于理解认知的概念工具,如主动推理[131,132],已经被用于理解形态发生和体内细胞能力的规模缩放[52,133]。
未来很可能涉及主流的分子药理学、计算推导出的利用现有药物的新方法[25,26],以及一种新的“身心医学”[62,134]的紧密结合。现有的计算工具已经可以预测治疗范式,以解决患者中的药物抗性问题,将其视为一种习惯化的形式,而不是开发新药物或靶向基因[25,26]。现在清楚的是,解剖结构,就像生理状态一样,并不是静态的——它需要积极维护[28,135–137]。成熟的组织结构在去神经化后会失去组织性,指向持续的信息维护。识别这些机制并学会与它们合作(除了微观管理细节)可能最终有助于解决不仅是大规模损伤和致癌转化的修复问题,还有衰老本身。我们认为,控制论、控制理论和行为神经科学可以为生物医学提供一个全新的工具包,通过针对组织层次的主体的感知、决策、记忆、问题解决能力、设定点和偏好,而不是分子和生理层次的身体适应性景观,来增进健康寿命。行为和适应性包括许多有用的科学概念框架,包括主动推理(以及一般的感知控制理论)、动态学习和表示、系统决策和资源优先级的第一人称视角、在稳态和适应态回路中的目标驱动行为,以及整合信息等。这些概念策略的定义,与基本单元性质无关,很容易通过信息等抽象视角,移植到非神经体细胞背景的生物医学中。这些策略不会将生物简化为硬连线机制,而是利用它们作为参与改善健康这一共同任务的活性主体的合作。在再生医学中实施这些策略的工具,类似于行为科学中用于与各种主体进行交流,并因此可预测地影响单个细胞到整个人类的工具。具体而言,可以利用脉冲药物刺激、光遗传学和声学来重置目标状态(解剖设定点)、训练细胞将期望的复杂响应与简单触发刺激相关联、重写当前状态信息(暴露于应激因素的组织的信息先验)、控制器官级别的应激反应、改变转录和解剖决策中的能量景观,并重新绘制功能模块的边界(通过控制组织区域中的缝隙连接边界)[138]。
当前最成功的生物医学干预措施,如药物和手术,针对的是体内的“入侵者”:微生物(抗生素)、寄生虫(抗寄生虫药物)和癌症等细胞缺陷(化疗)。这些干预措施能够实现真正的治愈——在干预结束后,稳定地恢复健康。相比之下,能够明确和可靠地引发宿主生物体结构和功能的永久修复的治疗方法很少。相反,大多数现有的药物干预措施只是针对症状,当停止药物治疗时,症状会再次出现。我们如何学会将生理回路推向稳定、健康的状态,并克服药物抗性和副作用的限制?
当前的医学方法有两个主要极端:通过药物作用于分子靶点来治疗症状,以及在各种手术后等待身体“康复”。相反,我们建议我们需要开发针对康复系统的干预措施,通过在体内读取和编写生理记忆来指定其目标状态。我们还应该努力增强它们的能力,就像使用认知增强剂(智力药物)一样(图7A)。
细菌[139]和寄生虫[140–142]已经在生物学的更高层次上进行了长时间的“黑客攻击”,我们也可以通过认识到细胞和组织是具有行动能力的材料[143],从而获得许多强大的预测和控制方法,这些方法在当前的机械性方法中被忽视了(图7B)。我们知道身体使用原型认知结构及其所带来的优势,但我们只是初步了解了每个与医学相关的子系统所提供的能力、目标状态和可行性。就像用行为科学中研究一种新生物一样[144],我们需要了解人类身体具有多少以及何种类型的行为可塑性,以及其能力和动机。然而,类似于动物训练,我们并不需要知道实现行为的所有机械细节才能控制它。
身体的历程,从根本上讲,是心智从物质中涌现的过程。我们每个人都曾经是一个静止的卵母细胞——一个化学物质团块,变成了复杂的人类元认知意识。在生命的黎明时期,使细胞能够解决生理问题的机制被转变为在发育过程中解决形态发生问题的机制,然后通过神经系统解决行为问题[57]。因此,关于问题解决能力的洞察,可以在神经认知科学和体内生理健康与疾病的分子生理学之间双向流动[63]。与通路、细胞、器官子系统和人体微生物组[13,145]进行沟通的手段将是生物医学进展的核心(图7C)。因此,未来的医学将更像精神病学,而不是化学(尽管通过新颖的生物工程界面[17,146,147]进行交付)。通过认真对待进化和发育生物学的连续性,并广泛借鉴信息和控制科学以及化学和生物物理学,我们可以实现在健康和增强人类能力方面的真正变革性前景。
1. Bugaj, L.J. et al. (2017) Interrogating cellular perception and de- cision making with optogenetic tools. J. Cell Biol. 216, 25–28
2. Lobo, D. et al. (2014) A linear-encoding model explains the var- iability of the target morphology in regeneration. J. R. Soc. Inter- face 11, 20130918
3. Baluška, F. et al. (2022) Cellular sentience as the primary source of biological order and evolution. Biosystems 218, 104694
4. Baluška, F. et al. (2022) Cellular and evolutionary perspectives on organismal cognition: from unicellular to multicellular organisms. Biol. J. Linn. Soc. 2022, blac005
5. Reber, A.S. and Baluška, F. (2021) Cognition in some surprising places. Biochem. Biophys. Res. Commun. 564, 150–157
6. Levin, M. (2023) Collective intelligence of morphogenesis as a teleonomic process. In Evolution ‘On Purpose’: Teleonomy in Living Systems (Corning, P.A. et al., eds), pp. 175–198, MIT Press
7. Csermely, P. et al. (2020) Learning of signaling networks: molecular mechanisms. Trends Biochem. Sci. 45, 284–294
8. Antebi, Y.E. et al. (2017) Combinatorial signal perception in the BMP pathway. Cell 170, 1184–1196
9. Mitchell, A. and Lim, W. (2016) Cellular perception and misperception: internal models for decision-making shaped by evolutionary experience. Bioessays 38, 845–849
10. Wilson, M.Z. et al. (2017) Tracing information flow from Erk to target gene induction reveals mechanisms of dynamic and combinatorial control. Mol. Cell 67, 757–769
11. Tweedy, L. et al. (2020) Seeing around corners: cells solve mazes and respond at a distance using attractant breakdown. Science 369
12. Tweedy, L. and Insall, R.H. (2020) Self-generated gradients yield exceptionally robust steering cues. Front. Cell Dev. Biol. 8, 133
13. Levin, M. (2019) The computational boundary of a ‘self’: developmental bioelectricity drives multicellularity and scale-free cognition. Front. Psychol. 10, 2688
14. Levin, M. (2022) Technological approach to mind everywhere:an experimentally-grounded framework for understanding diverse bodies and minds. Front. Syst. Neurosci. 16, 768201
15. Price, J. and Allen, S. (2004) Exploring the mechanisms regulat- ing regeneration of deer antlers. Philos. Trans. R. Soc. Lond.Ser. B Biol. Sci. 359, 809–822
16. Racovita, A. et al. (2022) Engineered gene circuits capable of reinforcement learning allow bacteria to master gameplaying. BioRxiv Published online June 21, 2022. https://doi.org/10. 1101/2022.04.22.489191
17. Clawson, W.P. and Levin, M. (2022) Endless forms most beau- tiful 2.0: teleonomy and the bioengineering of chimaeric and synthetic organisms. Biol. J. Linn. Soc. 2022, blac073
18. Blackiston, D. et al. (2021) A cellular platform for the develop- ment of synthetic living machines. Science. Robotics 6, eabf1571
19. Kriegman, S. et al. (2020) A scalable pipeline for designing reconfigurable organisms. Proc. Natl. Acad. Sci. U. S. A. 117, 1853–1859
20. Kriegman, S. et al. (2021) Kinematic self-replication in reconfigurable organisms. Proc. Natl. Acad. Sci. U. S. A. 118
21. Gumuskaya, G. et al. (2023) Motile living biobots self-construct from adult human somatic progenitor seed cells. BioRxiv Pub- lished online February 22, 2023. https://doi.org/10.1101/ 2022.08.04.502707
22. Emmons-Bell, M. et al. (2019) Regenerative adaptation to elec- trochemical perturbation in planaria: a molecular analysis of physiological plasticity. iScience 22, 147–165
23. Baluška, F. and Levin, M. (2016) On having no head: cognition throughout biological systems. Front. Psychol. 7, 902
24. Watson, R.A. et al. (2010) Associative memory in gene regula- tion networks. In Artificial Life XII, Proceedings of the Tenth In- ternational Conference on the Simulation and Synthesis of Living Systems (Fellerman, H. et al., eds), pp. 194–201, MIT Press
25. Biswas, S. et al. (2021) Gene regulatory networks exhibit sev- eral kinds of memory: quantification of memory in biological and random transcriptional networks. iScience 24, 102131
26. Biswas, S. et al. (2022) Learning in transcriptional network models: computational discovery of pathway-level memory and effective interventions. Int. J. Mol. Sci. 24, 285
27. Freddolino, P.L. et al. (2018) Stochastic tuning of gene expres- sion enables cellular adaptation in the absence of pre-existing regulatory circuitry. Elife 7, e31867
28. Rubin, H. (2006) What keeps cells in tissues behaving normally in the face of myriad mutations? BioEssays 28, 515–524
29. Nagato, T. et al. (1995) Effect of denervation on morphogenesis of the rat fungiform papilla. Acta Anat. (Basel) 153,301–309
30. McDowell, G. et al. (2016) From cytoskeletal dynamics to organ asymmetry: a nonlinear, regulative pathway underlies left-right patterning. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 371, 20150409
31. Xiao, J. et al. (2020) Epidemiological realities of alcoholic liver disease: global burden, research trends, and therapeutic prom- ise. Gene Expr. 20, 105–118
32. Delgado-Coello, B. (2021) Liver regeneration observed across the different classes of vertebrates from an evolutionary per- spective. Heliyon 7, e06449
33. Michalopoulos, G.K. (2017) Hepatostat: liver regeneration and normal liver tissue maintenance. Hepatology 65, 1384–1392
34. Vogel, A. et al. (2004) Chronic liver disease in murine hereditary tyrosinemia type 1 induces resistance to cell death. Hepatology 39, 433–443
35. Michalopoulos, G.K. and Bhushan, B. (2021) Liver regenera- tion: biological and pathological mechanisms and implications. Nat. Rev. Gastroenterol. Hepatol. 18, 40–55
36. Miyaoka, Y. and Miyajima, A. (2013) To divide or not to divide: revisiting liver regeneration. Cell Div 8, 8
37. Iansante, V. et al. (2018) Human hepatocyte transplantation for liver disease: current status and future perspectives. Pediatr. Res. 83, 232–240
38. Rhim, J.A. et al. (1994) Replacement of diseased mouse liver by hepatic cell transplantation. Science 263, 1149–1152
39. Nakamura, K. et al. (2007) Animal models of tyrosinemia. J. Nutr. 137, 1556S–1560S
40. Faraj, W. et al. (2010) Auxiliary liver transplantation for acute liver failure in children. Ann. Surg. 251, 351–356
41. McKiernan, P. (2013) Liver transplantation and cell therapies for inborn errors of metabolism. J. Inherit. Metab. Dis. 36, 675–680
42. Shanmugam, N.P. et al. (2011) Auxiliary liver transplantation: a form of gene therapy in selective metabolic disorders. J. Clin. Exp. Hepatol. 1, 118–120
43. Rela, M. et al. (1999) Auxiliary partial orthotopic liver transplantation for Crigler–Najjar syndrome type I. Ann. Surg. 229, 565–569
44. Burdelski, M. and Rogiers, X. (1999) Liver transplantation in metabolic disorders. Acta Gastroenterol. Belg. 62, 300–305
45. Dokmak, S. et al. (2013) Auxiliary liver transplantation with a small deceased liver graft for cirrhotic liver complicated by hepatocellular carcinoma. Transpl. Int. 26, e102–e104
46. Ren, W. et al. (2012) Integrating repopulation and regeneration of the auxiliarily transplanted small liver graft: the solution for organ shortage and immunosuppression. Med. Hypotheses 79, 241–245
47. Hoppo, T. et al. (2011) Rescue of lethal hepatic failure by hepatized lymph nodes in mice. Gastroenterology 140, 656–666
48. Komori, J. et al. (2012) The mouse lymph node as an ectopic transplantation site for multiple tissues. Nat. Biotechnol. 30, 976–983
49. Han, B. et al. (2022) Fat-associated lymphoid clusters as expandable niches for ectopic liver development. Hepatology 76, 357–371
50. Nicolas, C.T. et al. (2020) Ex vivo cell therapy by ectopic hepatocyte transplantation treats the porcine tyrosinemia model of acute liver failure. Mol. Ther. Methods Clin. Dev. 18, 738–750
51. Fontes, P. et al. (2020) Development of ectopic livers by hepatocyte transplantation into swine lymph nodes. Liver Transpl. 26, 1629–1643
52. Kuchling, F. et al. (2020) Morphogenesis as Bayesian inference: a variational approach to pattern formation and control in complex biological systems. Phys Life Rev 33, 88–108
53. Calvo, P. and Friston, K. (2017) Predicting green: really radical (plant) predictive processing. J. R. Soc. Interface 14, 20170096
54. Pezzulo, G. and Levin, M. (2016) Top-down models in biology: explanation and control of complex living systems above the molecular level. J. R. Soc. Interface 13, 20160555
55. Kramer, B.A. et al. (2022) Multimodal perception links cellular state to decision-making in single cells. Science 377, 642–648
56. Toettcher, J.E. et al. (2013) Using optogenetics to interrogate the dynamic control of signal transmission by the Ras/Erk module. Cell 155, 1422–1434
57. Fields, C. and Levin, M. (2022) Competency in navigating arbitrary spaces as an invariant for analyzing cognition in diverse embodiments. Entropy (Basel) 24
58. Boyle, E.A. et al. (2017) An expanded view of complex traits: from polygenic to omnigenic. Cell 169, 1177–1186
59. Kolodkin, A. et al. (2012) Understanding complexity in neurodegenerative diseases: in silico reconstruction of emergence. Front. Physiol. 3, 291
60. Waliszewski, P. et al. (1998) On the holistic approach in cellular and cancer biology: nonlinearity, complexity, and quasi-determinism of the dynamic cellular network. J. Surg. Oncol. 68, 70–78
61. Mathews, J. and Levin, M. (2018) The body electric 2.0: recent advances in developmental bioelectricity for regenerative and synthetic bioengineering. Curr. Opin. Biotechnol. 52, 134–144
62. Pio-Lopez, L. et al. (2022) Active inference, morphogenesis, and computational psychiatry. Front. Comput. Neurosci. 16, 988977
63. Pezzulo, G. and Levin, M. (2015) Re-membering the body: applications of computational neuroscience to the top-down control of regeneration of limbs and other complex organs. Integr. Biol. (Camb) 7, 1487–1517
64. Koshland, D.E. (1983) The bacterium as a model neuron. Trends Neurosci. 6, 133–137
65. Morimoto, B.H. and Koshland, D.E.J. (1991) Short-term and long-term memory in single cells. FASEB J. 5, 2061–2067
66. Yang, C.Y. et al. (2020) Encoding membrane-potential-based memory within a microbial community. Cell Syst. 10, 417–423
67. Prindle, A. et al. (2015) Ion channels enable electrical communication in bacterial communities. Nature 527, 59–63
68. Mathews, J. and Levin, M. (2017) Gap junctional signaling in pattern regulation: physiological network connectivity instructs growth and form. Dev. Neurobiol. 77, 643–673
69. Levin, M. and Martyniuk, C.J. (2018) The bioelectric code: an ancient computational medium for dynamic control of growth and form. Biosystems 164, 76–93
70. Fields, C. et al. (2020) Morphological coordination: a common ancestral function unifying neural and non-neural signaling. Physiology 35, 16–30
71. Vandenberg, L.N. et al. (2011) V-ATPase-dependent ectodermal voltage and pH regionalization are required for craniofacial morphogenesis. Dev. Dyn. 240, 1889–1904
72. Levin, M. (2021) Bioelectric signaling: reprogrammable circuits underlying embryogenesis, regeneration, and cancer. Cell 184, 1971–1989
73. Harris, M.P. (2021) Bioelectric signaling as a unique regulator of development and regeneration. Development 148, dev180794
74. Bates, E. (2015) Ion channels in development and cancer. Annu. Rev. Cell Dev. Biol. 31, 231–247
75. Zhao, S. et al. (2020) Biomedical applications of electrical stimulation. Cell. Mol. Life Sci. 77, 2681–2699
76. Reid, B. and Zhao, M. (2014) The electrical response to injury: molecular mechanisms and wound healing. Adv. Wound Care (New Rochelle) 3, 184–201
77. Zhao, M. et al. (2012) Electrical signaling in control of ocular cell behaviors. Prog. Retin. Eye Res. 31, 65–88
78. Tseng, A. and Levin, M. (2013) Cracking the bioelectric code: probing endogenous ionic controls of pattern formation. Commun. Integr. Biol. 6, 1–8
79. Mathews, J. et al. (2022) Ion channel drugs suppress cancer phenotype in NG108-15 and U87 cells: toward novel electroceuticals for glioblastoma. Cancers (Basel) 14
80. Churchill, C.D.M. et al. (2018) EDEn – Electroceutical Design Environment: an ion channel database with small molecule modulators and tissue expression information. iScience 11, 42–56
81. Pai, V.P. and Levin, M. (2022) HCN2 channel-induced rescue of brain, eye, heart and gut teratogenesis caused by nicotine, ethanol and aberrant notch signalling. Wound Repair Regen. 30, 681–706
82. Pai, V.P. et al. (2018) HCN2 Rescues brain defects by enforcing endogenous voltage pre-patterns. Nat. Commun. 9, 998
83. Pai, V.P. et al. (2015) Endogenous gradients of resting potential instructively pattern embryonic neural tissue via Notch signaling and regulation of proliferation. J. Neurosci. Res. 35, 4366–4385
84. Chernet, B.T. and Levin, M. (2013) Transmembrane voltage potential is an essential cellular parameter for the detection and control of tumor development in a Xenopus model. Dis. Models Mech. 6, 595–607
85. Chernet, B.T. et al. (2016) Use of genetically encoded, light-gated ion translocators to control tumorigenesis. Oncotarget 7, 19575–19588
86. Chernet, B.T. and Levin, M. (2014) Transmembrane voltage potential of somatic cells controls oncogene-mediated tumorigenesis at long-range. Oncotarget 5, 3287–3306
87. Chernet, B.T. and Levin, M. (2013) Endogenous voltage potentials and the microenvironment: bioelectric signals that reveal, induce and normalize cancer. J. Clin. Exp. Oncol. Suppl 1, S1-002
88. Levin, M. (2021) Bioelectrical approaches to cancer as a problem of the scaling of the cellular self. Prog. Biophys. Mol. Biol. 165, 102–113
89. Goel, P. and Mehta, A. (2013) Learning theories reveal loss of pancreatic electrical connectivity in diabetes as an adaptive response. PLoS One 8, e70366
90. Tseng, A.S. et al. (2010) Induction of vertebrate regeneration by a transient sodium current. J. Neurosci. 30, 13192–13200
91. Oviedo, N.J. and Beane, W.S. (2009) Regeneration: the origin of cancer or a possible cure? Semin. Cell Dev. Biol. 20, 557–564
92. Sahu, S. et al. (2017) Secrets from immortal worms: what can we learn about biological ageing from the planarian model system? Semin. Cell Dev. Biol. 70, 108–121
93. Shreesha, L. and Levin, M. (2023) Cellular competency during development alters evolutionary dynamics in an artificial embryogeny model. Entropy 25, 131
94. Tlsty, T.D. and Hein, P.W. (2001) Know thy neighbor: stromal cells can contribute oncogenic signals. Curr. Opin. Genet. Dev. 11, 54–59
95. Maffini, M.V. et al. (2005) Stromal regulation of neoplastic development: age-dependent normalization of neoplastic mammary cells by mammary stroma. Am. J. Pathol. 167, 1405–1410
96. Telerman, A. et al. (2010) Tumor reversion holds promise. Oncotarget 1, 233–234
97. Blackiston, D.J. and Levin, M. (2013) Ectopic eyes outside the head in Xenopus tadpoles provide sensory data for light-mediated learning. J. Exp. Biol. 216, 1031–1040
98. Blackiston, D.J. et al. (2017) Serotonergic stimulation induces nerve growth and promotes visual learning via posterior eye grafts in a vertebrate model of induced sensory plasticity. npj. Regen. Med. 2, 8
99. Murugan, N.J. et al. (2022) Acute multidrug delivery via a wearable bioreactor facilitates long-term limb regeneration and functional recovery in adult Xenopus laevis. Sci. Adv. 8, eabj2164
100. Ozugur, S. et al. (2022) Transcardial injection and vascular distribution of microalgae in Xenopus laevis as means to supply the brain with photosynthetic oxygen. STAR Protoc. 3, 101250
101. Ozugur, S. et al. (2021) Green oxygen power plants in the brain rescue neuronal activity. iScience 24, 103158
102. Magisetty, R. and Park, S.M. (2022) New era of electroceuticals: clinically driven smart implantable electronic devices moving towards precision therapy. Micromachines (Basel) 13, 161
103. Tan, T.H. et al. (2022) Odd dynamics of living chiral crystals. Nature 607, 287–293
104. Chao, Z.C. et al. (2008) Shaping embodied neural networks for adaptive goal-directed behavior. PLoS Comput. Biol. 4, e1000042
105. Kagan, B.J. et al. (2022) In vitro neurons learn and exhibit sentience when embodied in a simulated game-world. Neuron 110, 3952–3969
106. Mehrali, M. et al. (2018) Blending electronics with the human body: a pathway toward a cybernetic future. Adv. Sci. (Weinh) 5, 1700931
107. Staufer, O. et al. (2016) Functional fusion of living systems with synthetic electrode interfaces. Beilstein J. Nanotechnol. 7, 296–301
108. Levin, M. et al. (2020) Applications and ethics of computer-designed organisms. Nat. Rev. Mol. Cell Biol. 21, 655–656
109. Heyd, D. (2012) Is there anything unique in the ethics of synthetic biology? Perspect. Biol. Med. 55, 581–589
110. Evers, A.W.M. et al. (2018) Implications of placebo and nocebo effects for clinical practice: expert consensus. Psychother. Psychosom. 87, 204–210
111. Piedimonte, A. and Benedetti, F. (2016) Words and drugs: same mechanisms of action? J. Contemp. Psychother. 46, 159–166
112. Lui, F. et al. (2010) Neural bases of conditioned placebo analgesia. Pain 151, 816–824
113. Benedetti, F. et al. (2007) When words are painful: unraveling the mechanisms of the nocebo effect. Neuroscience 147, 260–271
114. Saatcioglu, F. (2013) Regulation of gene expression by yoga, meditation and related practices: a review of recent studies. Asian J. Psychiatr. 6, 74–77
115. Agnati, L.F. et al. (2012) Aspects on the integrative actions of the brain from neural networks to ‘brain-body medicine’. J. Recept. Signal Transduct. Res. 32, 163–180
116. Taylor, A.G. et al. (2010) Top-down and bottom-up mechanisms in mind-body medicine: development of an integrative framework for psychophysiological research. Explore 6, 29–41
117. Lu, H.Y. et al. (2021) Multi-scale neural decoding and analysis. J. Neural Eng. 18, ac160f
118. Betzel, R.F. and Bassett, D.S. (2017) Multi-scale brain networks. Neuroimage 160, 73–83
119. Gershman, S.J. et al. (2021) Reconsidering the evidence for learning in single cells. Elife 10, e61907
120. Nilsonne, G. et al. (2011) Learning in a simple biological system: a pilot study of classical conditioning of human macrophages in vitro. Behav. Brain Funct. 7, 47
121. Zoghi, M. (2004) Cardiac memory: do the heart and the brain remember the same? J. Interv. Card. Electrophysiol. 11, 177–182
122. Rogers, M.P. et al. (1983) Conditioned immunosuppression? Am. J. Psychiatr. 140, 1110–1111
123. Rogers, M.P. et al. (1979) The influence of the psyche and the brain on immunity and disease susceptibility: a critical review. Psychosom. Med. 41, 147–164
124. Rogers, M.P. et al. (1976) Behaviorally conditioned immunosuppression: replication of a recent study. Psychosom. Med. 38, 447–451
125. Miller, N.E. (1978) Biofeedback and visceral learning. Annu. Rev. Psychol. 29, 373–404
126. Ongaro, G. and Kaptchuk, T.J. (2019) Symptom perception, placebo effects, and the Bayesian brain. Pain 160, 1–4
127. Beauregard, M. and O’Leary, D. (2008) Believing can make it so: the neuroscience of the placebo effect. Adv. Mind Body Med. 23, 14–18
128. Mason, A.A. (1952) A case of congenital ichthyosiform erythrodermia of Brocq treated by hypnosis. Br. Med. J. 2, 422–423
129. Mathews, J. et al. (2022) Cellular signaling pathways as plastic, proto-cognitive systems: implications for biomedicine. Patterns (NY) 4, 100737
130. Busse, S.M. et al. (2018) Cross-limb communication during Xenopus hindlimb regenerative response: non-local bioelectric injury signals. Development 145, dev164210
131. Friston, K. (2010) The free-energy principle: a unified brain theory? Nat. Rev. Neurosci. 11, 127–138
132. Badcock, P.B. et al. (2019) The hierarchically mechanistic mind: a free-energy formulation of the human psyche. Phys Life Rev 31, 104–121
133. Ramstead, M.J.D. et al. (2019) Variational ecology and the physics of sentient systems. Phys Life Rev 31, 188–205
134. Adams, R.A. et al. (2016) Computational psychiatry: towards a mathematically informed understanding of mental illness. J. Neurol. Neurosurg. Psychiatry 87, 53–63
135. Rubin, H. (2007) Ordered heterogeneity and its decline in cancer and aging. Adv. Cancer Res. 98, 117–147
136. Rubin, H. (1992) Mechanisms for enduring biological change. Am. J. Phys. 262, L111–L113
137. Rubin, H. (1990) On the nature of enduring modifications induced in cells and organisms. Am. J. Phys. 258, L19–L24
138. Mathews, J. et al. (2023) Cellular signaling pathways as plastic, proto-cognitive systems: implications for biomedicine. Patterns (NY) 4, 100737
139. Williams, K. et al. (2020) Regulation of axial and head patterning during planarian regeneration by a commensal bacterium. Mech. Dev. 163, 103614
140. Eberhard, W. et al. (2014) Zombie bugs? The fungus Purpureocillium cf. lilacinum may manipulate the behavior of its host bug Edessa rufomarginata. Mycologia 106, 1065–1072
141. Elya, C. et al. (2018) Robust manipulation of the behavior of Drosophila melanogaster by a fungal pathogen in the laboratory. Elife 7, e34414
142. Loreto, R.G. and Hughes, D.P. (2019) The metabolic alteration and apparent preservation of the zombie ant brain. J. Insect Physiol. 118, 103918
143. Davies, J. and Levin, M. (2023) Synthetic morphology via active and agential matter. Nat. Rev. Bioengineer. 1, 46–49
144. Abramson, C.I. and Levin, M. (2021) Behaviorist approaches to investigating memory and learning: a primer for synthetic biology and bioengineering. Commun. Integr. Biol. 14, 230–247
145. Strassmann, J.E. and Queller, D.C. (2010) The social organism: congresses, parties, and committees. Evolution 64, 605–616
146. Rotenberg, M.Y. and Tian, B.Z. (2018) Talking to cells: semiconductor nanomaterials at the cellular interface. Adv. Biosyst. 2, 1700242
147. Belwafi, K. et al. (2021) Embedded brain computer interface: state-of-the-art in research. Sensors (Basel) 21, 4293
148. Levin, M. (2023) Darwin’s agential materials: evolutionary implications of multiscale competency in developmental biology. Cell. Mol. Life Sci. 80, 142
149. Krist, K.T. et al. (2021) A simple theory for molecular chemotaxis driven by specific binding interactions. J. Chem. Phys. 155, 164902
150. McGregor, S. et al. (2012) Evolution of associative learning in chemical networks. PLoS Comput. Biol. 8, e1002739
151. Craddock, T.J. et al. (2012) Cytoskeletal signaling: is memory encoded in microtubule lattices by CaMKII phosphorylation? PLoS Comput. Biol. 8, e1002421
152. Metzcar, J. et al. (2023) A model of multicellular communication mediated through extracellular matrix microstructure. BioRxiv Published online February 3, 2023. https://doi.org/10.1101/2022.11.21.514608
153. Sarris, M. and Sixt, M. (2015) Navigating in tissue mazes: chemottractant interpretation in complex environments. Curr. Opin. Cell Biol. 36, 93–102
154. Little, G.E. et al. (2009) Specificity and plasticity of thalamocortical connections in Sema6A mutant mice. PLoS Biol. 7, e98
155. Levin, M. et al. (2017) Endogenous bioelectric signaling networks: exploiting voltage gradients for control of growth and form. Annu. Rev. Biomed. Eng. 19, 353–387
156. Fankhauser, G. (1945) Maintenance of normal structure in heteroploid salamander larvae, through compensation of changes in cell size by adjustment of cell number and cell shape. J. Exp. Zool. 100, 445–455
157. McEwen, B.S. (1998) Stress, adaptation, and disease. Allostasis and allostatic load. Ann. N. Y. Acad. Sci. 840, 33–44
158. Zimmer, C. et al. (2022) Information theory in vertebrate stress physiology. Trends Endocrinol. Metab. 33, 8–17
159. Tschantz, A. et al. (2022) Simulating homeostatic, allostatic and goal-directed forms of interoceptive control using active inference. Biol. Psychol. 169, 108266
160. Deans, C. (2021) Biological prescience: the role of anticipation in organismal processes. Front. Physiol. 12, 672457
161. Colditz, I.G. (2020) A consideration of physiological regulation from the perspective of Bayesian enactivism. Physiol. Behav. 214, 112758
162. Schulkin, J. and Sterling, P. (2019) Allostasis: a brain-centered, predictive mode of physiological regulation. Trends Neurosci. 42, 740–752
163. Oviedo, N.J. et al. (2003) Allometric scaling and proportion regulation in the freshwater planarian Schmidtea mediterranea. Dev. Dyn. 226, 326–333
164. Cooke, J. (1981) Scale of body pattern adjusts to available cell number in amphibian embryos. Nature 290, 775–778
165. Levin, M. et al. (2018) Planarian regeneration as a model of anatomical homeostasis: recent progress in biophysical and computational approaches. Semin. Cell Dev. Biol. 87, 125–144
166. Farinella-Ferruzza, N. (1956) The transformation of a tail into a limb after xenoplastic transformation. Experientia 15, 304–305
167. Pilling, O.A. et al. (2017) Insights into transgenerational epigenetics from studies of ciliates. Eur. J. Protistol. 61, 366–375
168. Fields, C. and Levin, M. (2017) Multiscale memory and bioelectric error correction in the cytoplasm–cytoskeleton–membrane system. Wiley Interdiscip. Rev. Syst. Biol. Med. 10, e1410
169. Oviedo, N.J. et al. (2010) Long-range neural and gap junction protein-mediated cues control polarity during planarian regeneration. Dev. Biol. 339, 188–199
170. Vandenberg, L.N. et al. (2012) Normalized shape and location of perturbed craniofacial structures in the Xenopus tadpole reveal an innate ability to achieve correct morphology. Dev. Dyn. 241, 863–878
171. Slijper, E.J. (1942) Biologic anatomical investigations on the bipedal gait and upright posture in mammals – with special reference to a little goat born without forelegs II. Proc. Kon. Ned. Akad. Van Wetensch. 45, 407–415
172. Kozo-Polyansky, B.M. (1924) Symbiogenesis: A New Principle of Evolution, Harvard University Press
173. Noble, D. (2022) Modern physiology vindicates Darwin’s dream. Exp. Physiol. 107, 1015–1028
174. Levin, M. (2023) Bioelectric networks: the cognitive glue enabling evolutionary scaling from physiology to mind. Anim. Cogn. Published online May 19, 2023. https://doi.org/10.1007/s10071-023-01780-3
175. Pio-Lopez, L. and Levin, M. (2023) Morphoceuticals: perspectives for discovery of drugs targeting anatomical control mechanisms in regenerative medicine, cancer and aging. Drug Discov. Today 28, 103585
176. Pai, V.P. et al. (2012) Transmembrane voltage potential controls embryonic eye patterning in Xenopus laevis. Development 139, 313–323

Michael Levin,塔夫茨大学发育与合成生物学家,Vannevar Bush 杰出教授,领导艾伦探索中心和塔夫茨再生与发育生物学中心。
学者主页:https://as.tufts.edu/biology/people/faculty/michael-levin

Eric Lagasse,匹兹堡大学病理学系副教授,McGowan 研究所癌症干细胞中心的主任。
学者主页:https://mirm-pitt.net/staff/eric-lagasse-pharmd-phd/
大模型与生物医学:
AI + Science第二季读书会启动
生物医学是一个复杂且富有挑战性的领域,涉及到大量的数据处理、模式识别、理论模型建构和实验验证等问题。AI基础模型的引入,使得我们能够从前所未有的角度去观察和理解这个领域的问题,加速科学研究的步伐,提高医疗服务的效率和效果。这种交叉领域的合作,标志着我们正在向科技与生物医学深度融合的新时代迈进,对于推动科学研究、优化医疗服务、促进人类健康有着深远的影响。
集智俱乐部联合西湖大学助理教授吴泰霖、斯坦福大学计算机科学系博士后研究员王瀚宸、博士研究生黄柯鑫、黄倩,华盛顿大学博士研究生屠鑫明,共同发起以“大模型与生物医学”为主题的读书会,共学共研相关文献,探讨基础模型在生物医学等科学领域的应用、影响和展望。读书会从2023年8月20日开始,每周日早上 9:00-11:00 线上举行,持续时间预计8周。欢迎对探索这个激动人心的前沿领域有兴趣的朋友报名参与。
详情请见:
大模型与生物医学:AI + Science第二季读书会启动
生命是什么?生命怎样起源?生命怎样演化?这些是对生命现象的本质追问,除了传统的生物学研究,如今有大量来自信息、物理、计算机领域的工具方法,正在揭开生命复杂性谜题。基于此,集智俱乐部策划“生命复杂性”系列读书会,自2020年11月5日至今,近百名在从事相关问题研究的老师同学或感兴趣的朋友参与。
了解读书会具体规则、报名读书会请点击下方文章:
生命复杂性系列读书会:从信息和物理视角探索生命的内在逻辑
点击“阅读原文”,报名读书会