从生命到人工生命:未来生命形态会是怎样?

封面:Long Ken
导语
生命是一个复杂而神秘的现象。费曼曾说:“What I can’t create, I do not understand”。如果要理解生命,最好的方式恐怕就是去创造生命。无论是古犹太的魔偶(Golem)、古希腊的塔罗斯、中国周朝的偃师,还是文学作品中的弗兰克斯坦和现代科幻小说,这种创造生命的冲动跨越不同文明,贯彻整体人类历史。而今天,这种创造生命的冲动不单是为了满足功能或生产力的需求,更在科技实践中展现其潜力,尤其是在人工生命和人工智能领域,为理解自身和我们在宇宙中存在的意义做出了重要贡献。
人工生命(Artificial Life, Alife)是一个新兴的跨学科研究领域,它致力于通过计算模型和物理模拟,探索生命的各种潜在形式。Alife不仅试图模拟和复制地球上已知的生命形式,还尝试创造全新的、可能存在的生命形式,从而扩展我们对“生命”这一概念的理解。这包括使用计算机模拟生命过程,研究自组织系统,以及开发能模拟自然选择和进化过程的软件和硬件系统。本文将在回顾Alife历史的基础上,介绍其最新的研究方向和实践进展。
1. 从生命到人工生命
1. 从生命到人工生命
传统生物学中,生命通常被认为是由DNA和蛋白质构成,具有新陈代谢、生殖发育、遗传演化等一系列性状的自然系统。直到1944年,物理学家薛定谔发表《生命是什么》(What Is Life?),生命才摆脱特定的生理组织结构依赖,被定义为一个能量-信息耦合的负熵系统。这种思想既回应了过去,也迎合了当下。将生命从纯粹的有机论拉回到了有机论和机械论之间,也为Alife的理论研究提供了可能。
就在同一时期,许多不同领域的科学家开始了对Alife的探索,这一领域的探索大致可以分为三次浪潮:
第一次(20世纪50-60年代):自复制探索期
在这个时期,科学家们主要关注自复制、自组织和进化等生命维持和产生结构的基本原理。涉及的领域包括控制论、图灵机、形态发生学、神经网络模型和遗传算法。尤其是1948年冯·诺依曼发表《自复制自动机理论》,奠定了人工生命学科基础。
这一次浪潮,科学家们将生命视为一种逻辑形式,主要强调信息处理的抽象模型。在实践上,挪威数学家尼尔斯·巴里切利(Nils Barricelli)在1953-1962年于早期的计算机IAS上,便使用了一维元胞自动机,模拟了一个可以无限进化的数字世界。其中甚至出现了数字共生体生命形式。他据此认为,基于竞争进化模型的达尔文理论不足以解释生命演化的全貌,共生和合作在生命演化中占有重要地位[1]。这次浪潮从模型和算法层面梳理了生命的基本特征,祛除了生命发生的迷思,为人工生命奠定了理论和实践基础。
前两次Alife所持的功能主义立场以及抽象模型模拟,有助于抓住生命普遍本质特征。正如兰顿所相信,生命是一种独立于载体的恰当形式,不同的物质可以实现相同的生命和智能功能[3]。因此,这两次浪潮下的主要产物是软件/虚拟/数字人工生命(Soft)。

然而,在Alife研究的发展过程中,质疑和反思始终伴随其左右。同期活跃的理论生物学家罗伯特·罗森(Robert Rosen),在1991年出版了代表作《生命本身》(Life Itself),认为生命体是一个“有效因果闭合”(closed to efficient causation)的(M,R)系统*[4]:即系统中驱动变化发生的”因”,本身就必须是这个系统的”果”。例如生命系统中酶就扮演了有效因果闭合的角色,它们催化代谢反应的发生又产生自身。这种循环因果结构使生命具有某种自主性和自生成能力,意味着生物体不完全是机器,无法完全通过图灵机模拟。
(M,R)系统:即代谢(Metabolism)和修复(Repair)。代谢由一组机制(记为f)完成从材料A到产物B的转化,修复则由一组机制(记为Φ)从代谢产物B合成代谢机制f本身。
与此同时,生物和认知科学家弗朗西斯科·瓦雷拉(Francisco Varela)也反对兰顿的观点[5]。他认为生物体的情境性和历史性不可还原,生物体与其环境的具身交互远比其潜在的逻辑或功能模式更重要。这些思考引出了第三次人工生命浪潮。
第三波(20世纪90年代后期至今):嵌入式演化期
这一时期的研究更加关注人工生命与现实环境的关系,强调生命在真实世界的嵌入性、具身性、交互性和涌现性。1972年和1991年,瓦雷拉分别和合作者提出自创生(Autopoiesis)和具身心智(Embodied Mind)思想,分别强调生命系统的自我生成能力,以及与环境具身交互而产生的动态、嵌入式的涌现过程。这些都为人工生命研究提供了新的理论视角。
例如,研究者试图在系统内部引入“向上的张力”(避免系统落入平衡态的能力和结构),构建自指演化和动力学[7],以及将环境和观察者(如注意力和操作)纳入到系统功能描述中;或者在实验室进行实验,推动其在自组织递归循环中不断突破和产生创新功能和行为,在一个生态系统中向更广阔的方向演化。
除了数字人工生命外,湿件(Wet)、硬件(Hard),以及混合人工生命(Hybrid),成为这一时期的代表。这包括分子-细胞层面的人工化学、合成生物学、自复制机器人等领域的探索,研究范围从人工生命物发展到人工生态系统和社会。即便是在数字人工生命的研究中,科学家的重点也放在数字具身形式[8]和数字生态上。

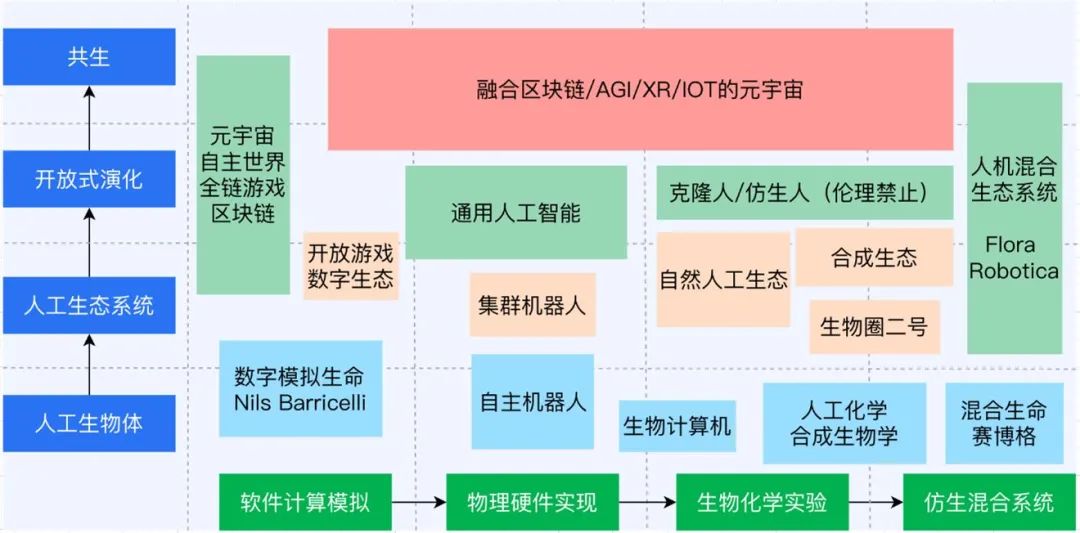
总得来看,我们可以把Alife发展分为两个轴,横轴是技术实现的方式,从软件计算模拟(软)→物理硬件实现(硬)→生物化学实验(湿)→仿生混合系统(混合),纵轴是研究焦点,从人工生物体 → 人工生态 → 开放式演化 → 共生(如下图)。
2. 人工生态与社会
2. 人工生态与社会
在自然界中,生物通常不是孤立存在的,例如,许多灵长类动物和社会性昆虫以种群的形式在生态系统中共生。生态系统,作为推动生命演化的核心力量,也由环境及其中的所有生物构成,表现为一个充满生态交互的动态系统。生态学的研究可以分为群落生态学和生态系统生态学,这两者分别关注生物种群间的互动及生物与物理环境之间的交互。与单纯创造人工生物体不同,这一分支的Alife研究的是相互作用的人工实体的集合。
在自然生态群落中,不同物种的生物相互作用。在Alife中,研究者则经常将个体视为某种基因型、表现型或生态型(genotypes,phenotypes,ecotypes)来分析群落动态。威廉·莱纳斯(William Reiners)[10]提出,统一的系统生态学研究需要至少三个独立互补的理论框架:能量学、物质学,以及种群相互作用或生态系统“连通性”(connectedness)的演化。这些框架可以分别理解为能量流、物质流和因果信息。
其中能量-物质的流动和转换,包括营养级、食物链和网[11]、生产率,地球生化学循环(如碳循环)等;种群互动或生态系统的连通性则包括种群之内的集体行为[12]、群落之间的相互作用(如捕食、竞争和共生)以及对生态位的研究,探索这些生态位如何涌现,消失,然后进化的[13]。
对ALife人工生态演化的研究,理论技术主要包括复杂动力系统、元胞自动机、反馈网络和控制论等,数字进化模拟平台则提供了检查复杂进化和生态系统动态的工具。
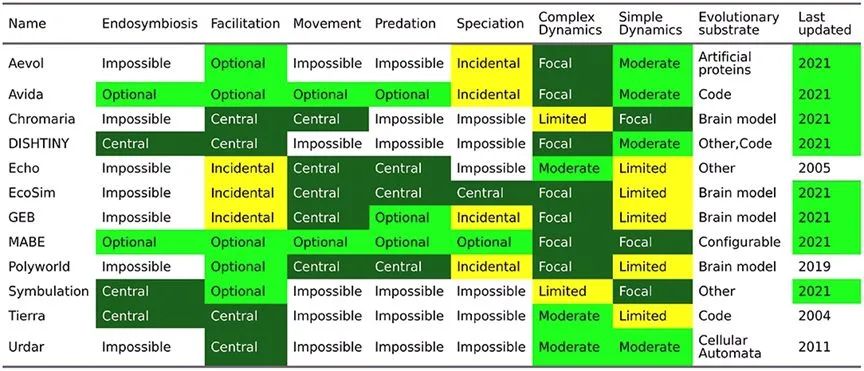
常用Alife数字进化模拟平台。[6]
其中一些模拟平台设置了能量-物质流模拟。例如,在Tierra中,能量被定义为执行指令所需的CPU周期(计算机处理时间)。智能体消耗CPU周期执行指令,改变其局部环境。因此,最小化能量消耗(CPU使用)可以提高复制效率,这为智能体施加了选择压力,促使其朝向更高效、可靠的演化方向发展。而在Avida平台中,生物体则通过代谢不同的有限资源(能量)相互竞争。
ALife 生态研究系统与自然系统不同之处在于,研究者可以设置远离自然界常见值的参数[15]。这样的设定就使得研究者能够探索哪些特征是生命所必需的,哪些特征和事件可能只是地球上生命形式的偶然产物。这样,ALife不仅能“重放前世的磁带”,还可以设计和播放与地球已存或可能运行非常不同的磁带(如火星),从而帮助澄清和提取生命的本质属性。
人工生态构建也分为软、硬、湿及混合人工生态四种形式。Alife数字平台能天然模拟软人工生态,其中还包括人工生命软件系统或游戏、及基于进化计算的数字生态艺术等。硬人工生命生态主要涉及机器集群系统的互动。湿人工生命生态则尝试构建通常不会共存的生物组成的生态群落,如执行理想任务(分解废物、封存碳等)的生态群落,这个子领域被称为合成生态学。至于混合人工生态,最著名的是 Flora Robotica 项目[16],该项目创建了一个植物和机器人共生的生态系统,机器人会通过在不同位置发射不同颜色的光来控制植物生长。

软件、湿件、硬件人工生态系统。
人工生态系统领域的研究和实践非常广阔。例如,随着复杂性的增加,生态系统可以促进社会和文化互动[17] 。其中涉及的掠夺、合作和文化的互动,引发了对感知、交流和语言的研究等[18]。此外,在人工智能领域,多智能体、人工集体智能的研究,也可以归结为人工生态系统和社会的研究[19]。
3. 开放式演化
3. 开放式演化
“除了演化的光辉,生物学中的一切都没有意义。”
——美国进化生物学家狄奥多西·多布赞斯基(Theodosius Dobzhansky)
在差异性选择的驱动下,进行繁殖、遗传和变异的生物种群和生态系统,会经历达尔文式的演化。进化算法[20]使用类似的概念来搜索外部环境适应度函数(fitness)的最优值,往往会带来令人意想不到的结果[21]。
然而,自然演化在多个层面上要比人工模拟复杂得多,包括基因组复杂性、种群规模、世代数量、从水平基因转移到性选择的繁殖策略、基因调控、发育和表观遗传学的作用、多个共同演化种群之间的互动以及内在定义的适应度等方面。这些更自然的演化特征构成了ALife演化研究的核心内容[14][22]。
如达尔文言,“无尽之形最美”[23],开放式演化(Open-Ended Evolution, OEE)[24],是ALife的一个重要研究领域。它不仅捕捉了现有生命系统各种属性、包括与环境作用关系的根本特征,同时提供了一个框架来探索和模拟潜在生命形式的可能性空间。在研究过程中,这有助于启发各种优化、学习和进化等算法。因此,实现开放式演化也可以说是人工生命的终极目标。
区别于一般演化,一个开放式演化系统永远不会陷入单一的稳定平衡,其新颖性会持续产生[25],复杂性会无限增加。这表明开放式演化是一个表征了多个尺度演化的过程[26]。开放式演化领域涉及生命起源问题[27]、复杂性和组织层次出现的重大演化转变[28]以及元演化,即演化能力及其演化的演化[29]等问题。
要产生开放式演化,有几个假设条件[30]:
(1)潜在基因型的无限遗传空间。这并不意味生物体的基因组长度必须没有限制地变化。由于主导基因的突变和遗传速度很慢,也可能存在大量非编码蛋白的调控基因,即原来所谓的垃圾基因,这些基因通过调控前者可以迅速产生多样性。这意味着只需要通过改变类似于神经网络中权重的调控开关,可能很快就能形成新的物种。这一假设可以帮助解释普遍共同祖先(LUCA)的生物共同基因,以及寒武纪生物大爆发。
(2)潜在表型之间存在多种突变途径。这意味着潜在生物体特征应该能够由许多不同的突变途径产生,例如人和章鱼的眼睛虽功能相似,但却是独立进化的;鱼和海豚的鳍足则是趋同演化的案例。理论生物学家斯图尔特·考夫曼(Stuart Kauffman)通过随机布尔网络对此进行了刻画,他发现基因调控网络的最终行为是由节点数量以及各节点的入度K决定的。当K=2时,网络就会进入既不是不动点、振荡 ,也不是完全混沌的混沌边缘的状态。这时网络每一种吸引子就对应一种细胞的基因表达,即不同的基因组,可以采取了不同的进化路径来达到与环境适配的同一目标表型。
(3)持续演化的动态自适应景观(Dynamic adaptive landscape)。这意味着生物体周围的环境是动态变化的。一方面,随着生物种群演变,其周围的环境也随其行为而改变,一方面动态的环境会不断对生物种群进行选择、通过表观遗传等作用影响基因型和表型的实现。
开放式演化可以在元胞自动机,数字进化模拟平台如Tierra、Polyworld、Avida等,人工化学[31]模拟平台Stringmol等[32]进行研究。2019年一篇研究[33],根据演化的表型行为与搜索空间的关系,确定了三种类型的新颖性以及对应的开放性:
探索开放性(Exploratory):可使用当前模型描述的新颖性,通常涉及现有组件的重新组合或现有参数值的修改。例如基因组中新的等位基因的组合可能决定了新脊椎动物的脊椎数目。
扩展开放性(Expansive):一种需要改变模型,但仍使用当前元模型中存在的概念的新颖性,涉及在状态空间中发现打开新邻域的状态。这往往涉及组织利用先前未被利用的化学或物理规律,或一个新的边界条件或装置的出现。例如飞行翅膀的出现,以及视觉感官系统的出现。
变革开放性(Transformational):一种引入新概念、需要改变元模型的新颖性。这不仅涉及利用新的物理定律,还包括了组织层级上的变迁。例如,合成一种以前未在代谢反应中使用的新化学物质。生命起源、真核细胞以及多细胞生物、大脑和自我意识、文字和技术都属于此类。这往往对应于复杂系统中的强涌现。
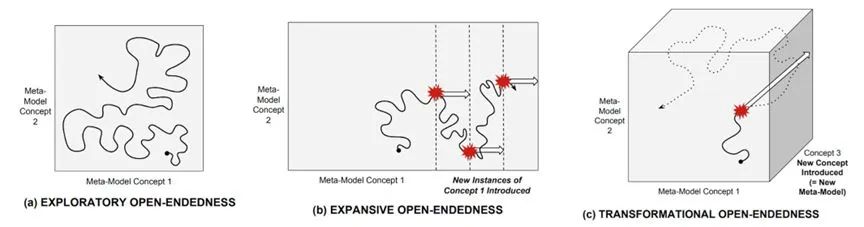
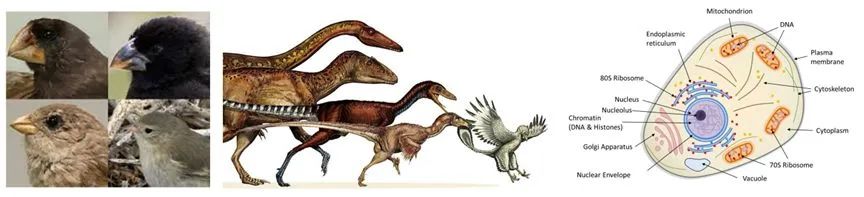
三种开放性:达尔文雀对生态位的适应;翅膀出现;真核细胞。
同样,对开放式演化有几种不同的度量指标也用于检测[34]或量化其开放性潜力,例如进化动态测量(MODES)指标[35]。在NK模型和Avida数字进化平台等不同的实验环境中,这些指标的有效性和适用性已经得到验证。
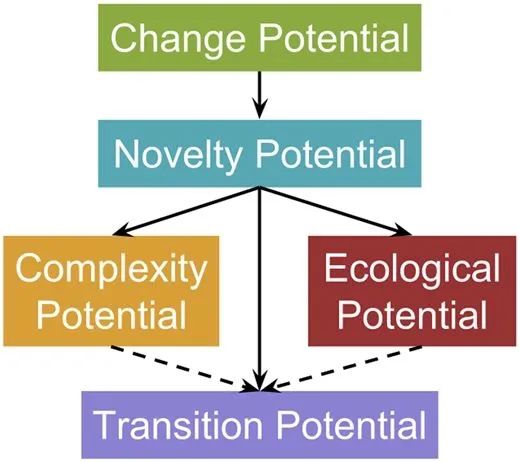
MODES各指标之间的关系。
在 MODES 中,复杂性潜力需要机体能够不断地将更多的环境信息整合到自身基因组中,从而产生更复杂的行为,生态性潜力描述的是种群作为一个整体能够吸收和反映环境信息的能力,包括生物与非生物环境相互作用创造新的生态位和营养层次的能力。
可见,要实现持续的开放性变革,必须将环境和生态因素纳入生物的演化过程中。据此,开放式演化系统可以视为Rosen的(M,R)系统。此外,据大语言模型ChatGPT描述,它更倾向于探索开放性。开放式演化的研究势必会对计算创造力和AGI研究提供重要的方法和范式。
4. 共生
4. 共生
“演化中的一切,只有在寄生的光芒下才有意义”,这篇2021年一篇论文的标题[36],模仿了多布赞斯基对演化的描述,但这或许并不夸张。诸多研究表明,寄生-互利共生连续谱的演化[37],是生成生物新颖性、塑造生态多样性和推动地球生命重大转变的重要机制。如前所述,尼尔斯·巴里切利的人工生命模拟早期就洞悉了这点。尼克·莱恩(Nick Lane)在《复杂生命的起源》中也认为,内共生提高了生命能量利用效率,促进了真核生物的起源,并使其最终演化成地球复杂生命。
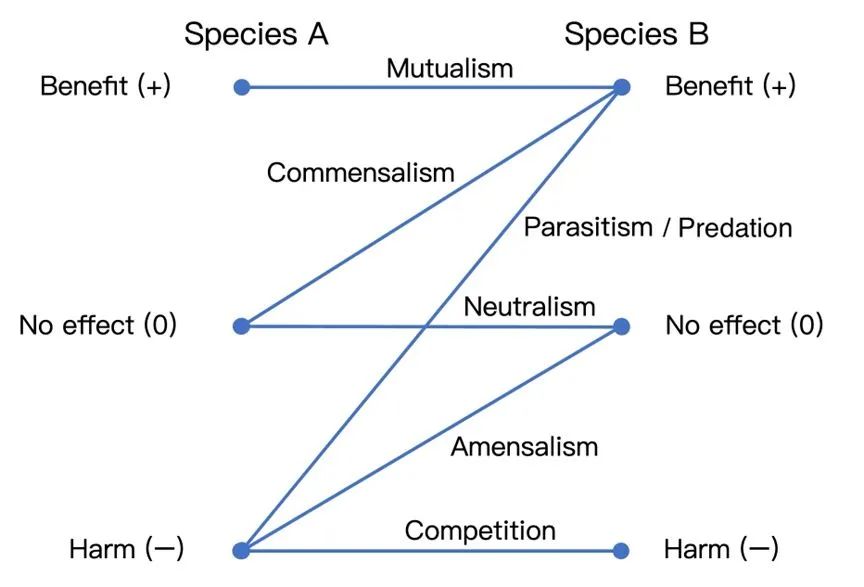
共生(Symbiosis)可以看做是生物体和种群之间,在更多样的生态尺度下更为密切的协同演化[38]的结果。根据空间位置,共生可以分为内共生(ectosymbiosis)和外共生(endosymbiosis),根据物种间的利益关系,则通常包括互利共生、偏利共生、中性互作、偏害共生、竞争以及寄生[39]/捕食(广义)[40]。生态系统中资源交换-利用依赖循环会导致社群构建和共生演化,甚至形成跨喂养(crossfeeding)现象,即一个物种的代谢物成为另一个物种的资源[41]。
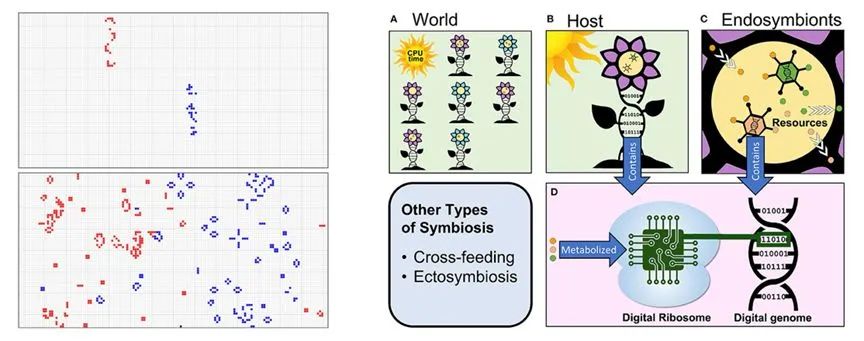
共生同样可以用生命游戏、数字进化模拟平台进行研究。例如,有研究者在康威的生命游戏基础上发展了Model-S模型[43],该模型成功模拟产生了自组织、自创生、多细胞的演化、有性生殖的诞生以及物种共生的融合策略。研究发现,即使是少量共生也能显著提高种群的适应度,并可能支持开放式演化。在支持共生的数字进化系统中[44],数字生物通过消耗“CPU时间”资源来生存和发展,内共生物则消耗宿主内部资源,与真实世界中细胞的线粒体类似。同样,数字核糖体将数字基因组(二进制代码)转化(代谢)为某种输出或行为,类似于生物细胞中核糖体读取遗传代码并合成蛋白质的过程。
可以说,生命从起源起就是一种共生现象,并且横跨了不同种类的复制子。从基因到文化(模因),到以技术作为扩展表型(技因),未来,生物与数字的融合可能进入一个人机共生的新时代[45]。
如果生物演化的基质基于自然物理环境,那么技术物-数码物的演化基质和生态环境又是什么?显然,这必须是一个人类和机器生命都能进行互操作的系统。有研究指出[46],基于区块链的分布式系统具有对环境响应、生长和变化、遗传复制和达到稳态等诸多符合生命定义的特征。若和神经网络等AI技术结合,这种系统将成为比传统生命更有优势的自组织系统。文章认为,基于公链的分布式虚拟机(dVM)提供了一个自主进化能力的开放环境,为通用人工智能(AGI)的发展提供支持。
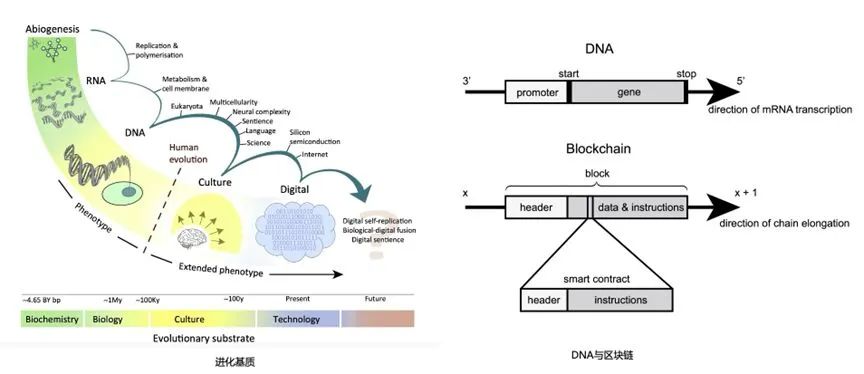
进化基质的演化[45];DNA与区块链[46]。
最近,区块链技术领域兴起了一种名为数字物质理论(Digital Matter Theory)的观点,该理论认为,数字信息可以被看作一种数字物质,与实体物质(比如木头或金属)类似。通过挖掘区块数据中固有的模式,可以创建一种新型的数字物质形态,即区块元素表。它被视为数字世界物质基质和资产的一种表现。随着区块高度增加,这个数字世界也和物质世界一样在不断生长,其发展同样不可预期。在这样的系统中,人们可以发行非任意同资源/代币(Non-Arbitrary,意味着总量非人为设定或随意增发),为自主世界与元宇宙提供了底层逻辑和基础。
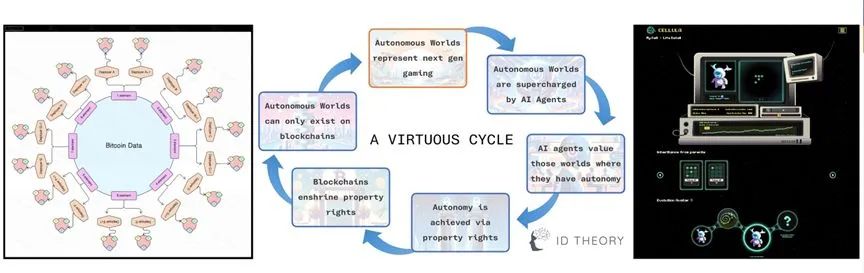
数字物质理论、自主世界和链上生命游戏Cellula[55]。
此外,目前也出现一些基于区块链的Alife项目。例如链上的人工生命模拟游戏(Cellula),它允许玩家自由地创建各种基因序列,并孕育出自己的链上“生命”,在这个游戏中,区块高度扮演了“时间”的角色,每个“生命”都会在链上时空生态系统中成长、进化、死亡。区块链上的Alife,由于涉及到了玩家和链上智能体主体,存在注意力流和信息的双向传递,因此是一种典型的互惠共生形态。此外,还有一些项目探索了通过区块链技术或物理环境,使非人类主体如植物与数字或机器共生的可能性,例如terra0和 Flora Robotica项目。

跨媒介跨物种共生的例子。
当然,比较广泛讨论的人机共生是人和人工智能方面和共生。这主要基于任务和协作的研究、以及人机价值对齐[47]、尤其AI心智理论[48]方面的研究。这些研究在很大程度上体现了人类中心主义的观点以及以人类利益为主的共生设计考量。
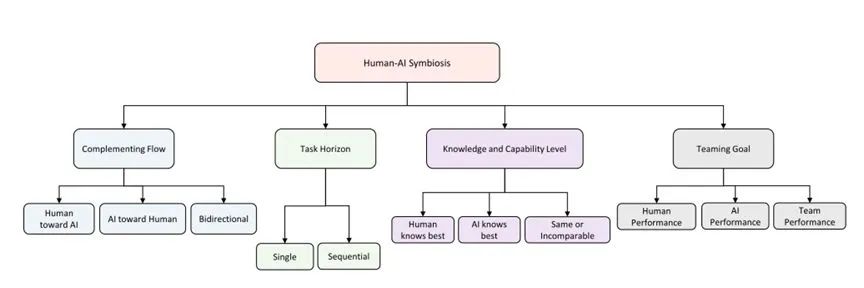
人机共生的方法维度。[49]
5. 生物启发AI与人工生命的未来
5. 生物启发AI与人工生命的未来
回顾历史可以看到,AI曾长期受制于符号人工智能桎梏,即计算启发方法(GOFAIstic)[50],而Alife和生物启发方法(亦被称为控制论启发)则为AI的发展提供了重要灵感和范式。
辛顿(Geoffrey Hinton)开创的深度学习网络,实际上是一种模拟人脑神经的AI系统,而像Transformer这样的大模型通过自注意力机制在某种程度上模拟了人类大脑的记忆和学习机制,即人脑的海马体记忆模型是一个带了循环神经网络的因果Transformer(“TEM is causal transformer with RNN position encodings”)[51]。开发出 Chatgpt 的OpenAI,时刻提醒员工“苦涩的教训”(The bitter lesson),坚信规模法则(Scaling Law)就能涌现属性(Emerging properties) ,认为只要持续不断增大数据和算力的大模型才能实现AGI。强化学习包括人类反馈强化学习(RLFH),源自对动物行为心理学的借鉴,本质是一种基于反馈的环境适应机制,生成式对抗网络(GAN)则源自动物竞争与共生的生态学机制。
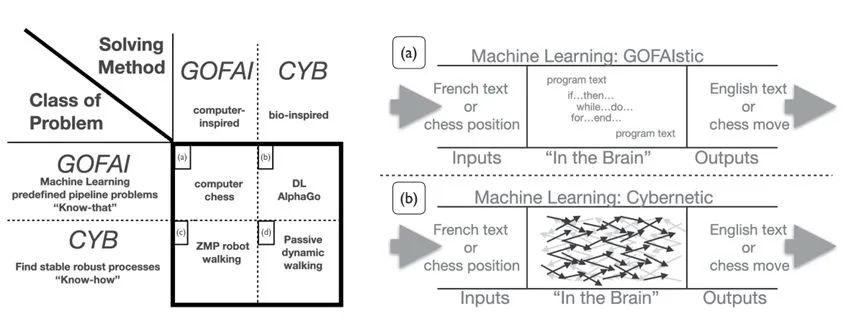
GOFAI主义与生物启发(控制论)AI[52]
随着技术进步,Alife研究者不断探索从抽象模型到计算模拟,再到具身嵌入、人工生态、开放式演化等多种范式,这些都在推动AI的进一步发展。例如,卡尔·弗里斯顿(Karl Friston)的自由能原理和杨立昆的联合嵌入预测架构(I-JEPA)[53]都尝试在AI系统中整合更多生物学原理,以增强其与环境的交互能力和自主性。甚至包括非冯诺依曼架构、辛顿后来提出的可朽计算(Mortal Computation)、如今广为讨论的人机价值对齐,以及受生态启发的人工智能[56]等研究,都可以看到Alife的发展进路的影子。
当然,目前的生成式AI,特别是大语言模型,也在对Alife研究产生深远影响[54]。例如利用大语言模型探索意义的产生、涌现和因果涌现、开放式演化,尤其是人机协作、人工集体智能以及大语言模型驱动的多AI智能体系统(著名的斯坦福AI小镇)。此外,目前也有算法层面的结合,例如结合神经网络和遗传算法的神经进化(Neuroevolution),已有研究者使用简单的虚拟生物[57],通过视觉、听觉感知环境,并与同类和捕食者互动,发现部分个体涌现出专门应对捕食者的”恐惧中枢”模块,这表明类似生物脑的情绪和意识等神经机制可以在人工基质中涌现。
最后,如果我们以Alife的范式去思考,我们面前展开的问题有:基于区块链的系统和自主世界等数字原生基质如何实现具有自主性的(M,R)系统?大语言模型和AI智能体如何演化出变革开放性?AI和人类将会是形成哪种共生关系,竞争、内共生还是跨物种喂养?虚拟现实和扩展现实又将如何影响未来的数字生态?
不管未来如何,Alife、AI、区块链和元宇宙(AR/VR/XR)将相互交织发展,带来一个自然生命和虚拟主体、数字和物理的相融的共生世界。在这样的后人类时代,不管是哪种形式,生命实体都将在广阔无垠的宇宙中继续其开放演化的旅程。
参考文献
(参考文献可上下滑动查看)


AI By Complexity读书会招募中
大模型、多模态、多智能体层出不穷,各种各样的神经网络变体在AI大舞台各显身手。复杂系统领域对于涌现、层级、鲁棒性、非线性、演化等问题的探索也在持续推进。而优秀的AI系统、创新性的神经网络,往往在一定程度上具备优秀复杂系统的特征。因此,发展中的复杂系统理论方法如何指导未来AI的设计,正在成为备受关注的问题。
集智俱乐部联合加利福尼亚大学圣迭戈分校助理教授尤亦庄、北京师范大学副教授刘宇、北京师范大学系统科学学院在读博士张章、牟牧云和在读硕士杨明哲、清华大学在读博士田洋共同发起「AI By Complexity」读书会,探究如何度量复杂系统的“好坏”?如何理解复杂系统的机制?这些理解是否可以启发我们设计更好的AI模型?在本质上帮助我们设计更好的AI系统。读书会于6月10日开始,每周一晚上20:00-22:00举办。欢迎从事相关领域研究、对AI+Complexity感兴趣的朋友们报名读书会交流!
推荐阅读
点击“阅读原文”,报名读书会
微信扫一扫,分享到朋友圈











