复杂生命的起源和兴起特刊:综合模型、地球化学和古生物学数据

导语
从原核生物到真核生物,从单细胞生物到多细胞生物,复杂生命如何起源,并在地球上繁荣昌盛?这个生命进化中的核心难题已经困扰我们数百年时间。从生物学角度来看,化石记录是进化史的唯一直接证据,但是真核生物的标志——细胞核和细胞器往往难以经历漫长地质年代保存下来,因此对复杂生命起源的研究可谓困难重重。不过近年来,通过整合来自地球化学、古生物学、古环境学、古地理学、基因组学等相关学科的丰富数据集,科学家对7.2-2.1亿年前复杂生命起源的具体时间和触发因素有了更深入的理解。
英国皇家学会曾于2019年9月举行了“复杂生命的起源和兴起:综合模型、地球化学和古生物学数据”讨论会,对气候变化、海洋氧化还原、营养利用率及生态系统反馈等一系列因素在生命起源期中所扮演的角色进行了跨学科的数据整合与假设检验。讨论中涉及到的15篇文章被整合收录至英国皇家学会旗下交叉学科期刊 Interface Focus 2020年8月刊。集智翻译了这个合集中1篇综述的部分内容以及14篇文章的摘要,供对复杂生命起源问题感兴趣的朋友参考。
研究领域:复杂生命起源和进化,地球化学,古生物学

Interface Focus | 来源
张澳 | 译者
徐恩峤、梁金 | 审校
邓一雪 | 编辑

链接:https://royalsocietypublishing.org/toc/rsfs/2020/10/4
目录
综述文章
复杂生命的起源和兴起:进步需要跨学科的整合和假设检验
14篇论文摘要
1.调和新元古代和古生代地球氧化的代用资料和模型
2.论模型在认识复杂生命起源中的应用
3.早期动物进化中的生存和选择偏差以及分子钟系统性高估的一个来源
4.从化石记录看真核生物
5.吞噬作用在地球历史上的起源
6.埃迪卡拉纪早期瓮安类胚胎化石中的细胞核保存、细胞核的实验化石形成学及对解释真核化石记录的启示
7.硅铝酸盐包层保存了大约8亿年前的复杂生命
8.内华达州埃迪卡拉纪森林峡谷地层中富含粘土的硅塑性沉积物中 erniettomorph 化石的保存
9. 动物的起源和早期进化:从遗迹化石的角度看我们处于什么位置?
10.埃迪卡拉纪的生物层积学:埃迪卡拉纪生物的沉积环境、生态学和生物学意义
11.环境条件对埃迪卡拉纪生物群落交错区的影响
12.利用缺氧条件作为复杂多细胞生物进化的驱动力
13.风化、变质和重建地球的氧化作用
复杂生命的起源和兴起:
进步需要跨学科的整合和假设检验
复杂生命的起源和兴起:
进步需要跨学科的整合和假设检验
The origin and rise of complex life: progress requires interdisciplinary integration and hypothesis testing
作者:Rachel Wood,Philip C. J. Donoghue,Timothy M. Lenton,Alexander G. Liu,Simon W. Poulton
发表时间:2020年6月12日
论文地址:https://royalsocietypublishing.org/doi/10.1098/rsfs.2020.0024
科学界对复杂生命起源和兴起的兴趣已有数百年的历史。自达尔文的《物种起源》[1]以来,它一直被视为进化论的核心难题之一。然而,尽管拥有地球化学、古生物学、古环境学、古地理学、基因组学等相关学科的丰富数据,但复杂生命起源的问题仍未得到较好解决,原因何在呢?
至少有两个显然的原因:一方面,跨学科数据难以整合;另一方面,生命起源相关假设可检验性较差。前者指来自各学科的数据不能被整合、校准至同一时空框架,因为对底层剖面的地球化学记录并未统一使用相同的地质年龄模型,这导致对同一生化循环或地球循环随时空变化的规律无法得到综合的理解。后者指在数据得到整合后,需要利用数据检验假设是否成立。然而,大多数生命起源的假设不够具体,难以证伪,部分假设甚至相互矛盾。除此之外,不少假设并不是针对某一特定的演化支(clade)提出的。比如,一些研究人员并未将后生动物的起源作为关键对象,而是聚焦于更复杂的真后生动物、两侧对称动物。实际上,无论真后生动物或两侧对称动物起源的原因是什么,都不会具有普遍性,因为生命起源不可能存在决定性的驱动因素。因此,需要明确生命的分类(最好具有解剖学及生态学证据),并将生命起源的因果机制与分类结果联系起来。
这些问题的有效解决需要从以下几个方面改进:(1)优化基础数据并接受其固有的不确定性;(2)更有效地整合不同时空框架下的数据集,区别局部模式与全局模式;(3)改进根据校正后的数据进行假设检验的模型。
上述建议源自2019年9月英国皇家学会组织的讨论会,它成功地激发了各学科研究者对生命起源中基本问题的讨论。我们相信这些挑战并非不可克服,只是需要借助领域的一致努力。该主题的研究证明我们对复杂生命起源和兴起的知识正以惊人的速度累积,我们希望这种势头能够持续下去,激发生命复杂性起源和兴起的问题取得真正的突破。
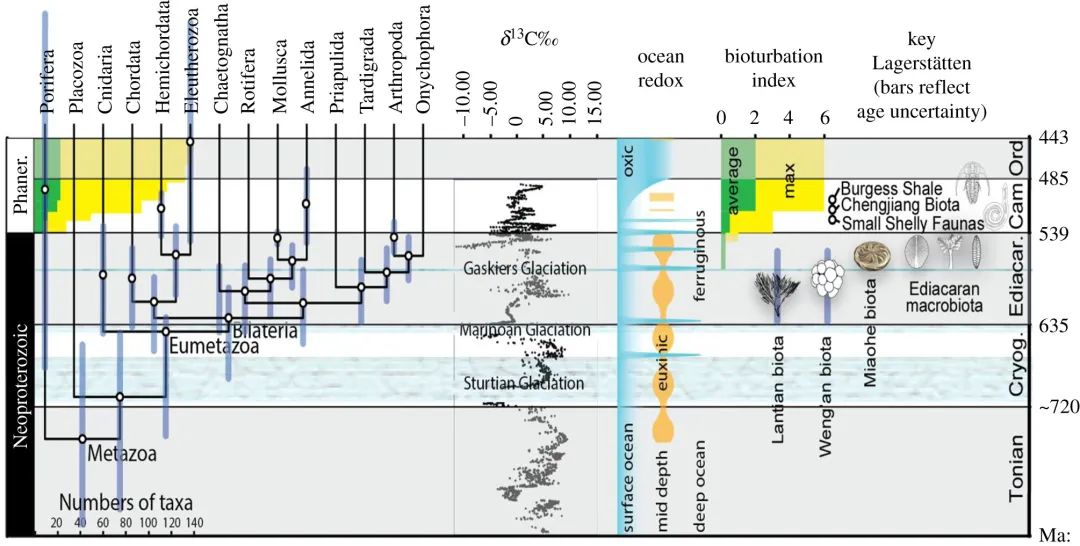
相关文献概述
以下是对这个主题下部分重要研究的总结,探索不确定性的问题,并检验围绕关键进化事件和具体主题的假设。其首先讨论了真核生物的起源,以及由于埋藏学(又称化石形成学,研究化石形成与保存规律的学科)的变幻莫测而产生的对化石记录的解释偏差;其次考虑了如何探寻并量化最古老的后生动物群落的生态动力学,以及氧气对早期动物生理上的挑战(而不是促进);随后思考了次生变化对地球化学信号的影响,以及如何选择适当的原则和概念框架以支持各模型的建立和检验;最后探讨了复杂生命的演变与地球系统在地质时间中的稳定性间可能存在的长期关系。
1. 真核生物的起源
Porter[2]提出了一个关于真核生物起源及演化的颠覆性假设,即真核生物起源于古元古代晚期,详见第4篇摘要。Mills[3] 进一步认为,真核细胞的吞噬作用(真核细胞将细胞膜包裹在其他物体周围并将其吞噬的能力)进化得很晚,而且是经过多次进化形成的,这与教科书上的观点相反,详见第5篇摘要。
2. 化石记录及其保存
为了认识到过去生物进化和全球环境之间的相互作用,我们需要对生物和环境的记录数据及其解释有信心。从生物学的角度来看,化石记录是进化史的唯一直接证据,但是从化石中剔除不需要的噪声并分离出真正的生物特征,对于我们准确解释某一特定保存特征具有生物系统发育信息的可能性至关重要。该主题概述了现有化石记录的主要方面。
解释早期真核生物的化石记录是具有挑战性的,因为它们具有区分度的主要特征(如细胞器和细胞核)通常不会被保存下来。即使确认了化石的存在,对其解释仍然存在高度争议。Sun等人[4]试图通过验证细胞核化石确实可以保存的假设,来减少真核生物出现时间的不确定性,详见第6篇摘要。
我们对有机真核生物微化石的早期记录大部分来自细密的、富含粘土的岩石(泥岩),这就引出了一个问题,为什么这样的环境有利于特殊的保存?这样的保存环境在空间或时间上是受限制的吗?利用高分辨率的原位地球化学技术,Anderson 等人[5]验证了某些矿物能够促进有机物石化的假说,详见第6篇摘要。他们发现,高岭石有促进化石保存的作用,有助于保存寒武纪伯吉斯页岩和类似沉积物的非骨骼后生动物[6]。先前研究表明,高岭石可以减缓细菌分解者的生长,并通过提供一种保护性的涂层[7]来保存软组织。另有研究表明,早期化石本身也可能促进了有机分子聚合和高岭石富集[8]。
Hall 等人[9]对来自内华达州埃迪卡拉纪(又称震旦纪)晚期的含粘土砂岩软体生物的大型化石采用了类似的分析方法,详见第8篇摘要。作者注意到化石中没有硫化物矿化或二氧化硅胶结物的证据,而这两者是一些埃迪卡拉纪晚期软体生物的化石形成学模型预期的矿物[10,11]。这就提出了关于化石保存过程的问题。正如 Anderson 等人[5]的研究,他们解释了粘土矿物在化石保存过程中的作用。这是否意味着粘土矿物是埃迪卡拉纪化石保存不可或缺的组成?
遗迹化石(trace fossil)记录了生物的行为,而不仅仅是它们的残骸。Mángano & Buatois [12]概述了埃迪卡拉纪-寒武纪过渡时期的两侧对称动物的痕迹,强调了生物体的活动能够实质性地改变它们所居住的生态系统的性质[13,14],详见第9篇摘要 。Mángano & Buatois 还提出了一个四阶段模型,概括了从埃迪卡拉纪晚期到寒武纪第四期两侧对称动物运动行为的变化。该模型除了阐明两侧对称动物运动行为进化的轨迹外,还澄清了关于寒武纪底质革命[15](Cambrian Substrate Revolution)和农艺革命(Agronomic Revolution)在时间上的脱节(看起来是用混合底质代替了藻席底质[16])。
3. 早期复合群落的生态学
关于埃迪卡拉纪深海大型生物群的生物多样性和结构的规则仍具有根本性的意义。该规则适用于控制现代生态系统的动力学吗?环境是如何影响生态的?位于 Nilpena 的埃迪卡拉化石好似一个自然实验室,提供了机会来确定生物层积学对埃迪卡拉纪大化石层理复合物产生的特定影响。Droser 等人[17]利用他们在挖掘多个广阔层理面露头[18](bedding plane surfaces)过程中获得的知识,提供了一个古生态基准,可以与被怀疑受到环境条件影响的露头进行比较,详见第10篇摘要。
一旦清楚地了解了化石形成学偏差对化石群的影响程度,就可以验证关于“组装规则(rules of assembly)”的假设。通过对7个埃迪卡拉纪晚期(558-550Ma,其中Ma指mega annum,代表一百万年,常用于描述地质年龄)大型化石组合的研究, Mitchell 等人[19]利用空间点过程分析来检验环境异质性可能控制化石群结构的假设,详见第11篇摘要。
Budd & Mann [20]根据动物的早期记录及其不存在于更古老地层中的证据,挑战对成冰纪(Cryogenian,又称南华纪)或拉伸纪(Tonian,又称青白口纪) 古遗迹基于分子钟的地质年代估计,详见第3篇摘要。
4. 早期动物面临的氧气问题
Hammarlund [21]对氧气在早期动物进化中的作用提出了新的看法。细胞分化是组织发育中的一个微妙过程,在该过程中,部分 DNA 无法转录,而其他 DNA 仍然可以转录,从而使细胞具有特定的命运。而氧气可凝结(condense)DNA(异染色质),限制部分基因的转录,使细胞分化。与分化所需的氧气条件不同,促进组织更新的干细胞在低氧甚至缺氧条件下工作。这就产生了悖论:有氧条件使组织分化,而组织的更新和维持则依赖于缺氧条件。本文结合干细胞研究、发育生物学及地质记录,提出大型(多样化)生命需要缺氧条件的假设,并给出了相应证据及该假说的影响。详见第12篇摘要。
5. 地球化学和模拟
地球化学提供了一个主要的平台,通过它可以探讨与早期动物进化有关的环境驱动因素和反馈。在过去20年中,在新技术和创新性假设的推动下,利用地球化学和同位素技术以重建动物生命初期环境条件的研究出现了爆炸式增长。所有这些研究的内在前提是,我们如今在岩石中测量的信号,是在沉积时期或成岩作用后最初被困在岩石中的信号。然而,随着大量研究的产生,很难控制这一前提,特别是对于氧化还原敏感的元素在沉积后极易发生价态的变化。
Planavsky 等人[22]坚定地把该问题放在研究的首位,根据当前学界在整个地球历史中的稀土元素主要数据集,详细研究了其次生变化(secondary alteration),详见第13篇摘要。
Tostevin & Mills [23]回顾了一个新元古代至古生代大气氧化还原状态和海洋氧化还原状态的代用资料,表明在埃迪卡拉纪-寒武纪期间,大气中的氧含量在0.1~1 PAL (目前的大气水平)之间波动,详见第1篇摘要。参考文献[24]说明了模型中出现这种情况的原因。
Tostevin & Mills [23]证明,将地球化学代用指标的数据与生物地球化学模型制约因素结合起来,为检验环境与各种形式复杂生命进化之间潜在联系的假说和反馈提供了巨大的可能性。然而,对于大部分研究者(包括重视上述方法的研究者)而言,他们意识不到自己在建模。对于模型的应用者而言,很难评估个别模型的局限性,或者难以评估模型对于不同问题的适用性。在一篇关于现有模型及其局限性和未来可能性的学术评论中,Lenton[25]为模型应用者打开了一扇通往建模世界的窗口。他的方法侧重于复杂生命的演变和多样化对氧气和营养水平重建的影响,更有价值的是,他概述了所有生物地球化学模型应遵循的原则,以及对现有关键模型的批判性评价,详见第2篇摘要。
Payne 等[26]考虑了显生宙(Phanerozoic)复杂生命的进化和地球系统表面上的稳定性之间的关系。显生宙的背景灭绝率在下降,但这是为什么呢?Payne等人认为,动物已经进化到对环境变化具有较高的生理弹性。此外,更大型的、移动的动物的进化[27]及水柱被认为已经稳定了生物地球化学循环。总的来说,Payne 等人同意并补充了先前的分析[28] ,即“复杂生命的进化总体上加强了气候系统中促进稳定的反馈”[26],详见第14篇摘要。这是一个非凡的发现,与 Lovelock 和 Margulis’ Gaia 的假说[29]一致,即随着时间推移,生命的进化一直在使地球系统更加稳定和有弹性。
参考文献
(参考文献可上下滑动查看)
以下为 Interface Focus 当期另外14篇论文(研究论文及评述文章)的摘要:
1. 调和新元古代和古生代地球氧化的代用资料和模型
论文题目:Reconciling proxy records and models of Earth’s oxygenation during the Neoproterozoic and Palaeozoic
作者:Rosalie Tostevin,Benjamin J. W. Mills
发表时间:2020年6月12日
论文链接:https://royalsocietypublishing.org/doi/10.1098/rsfs.2019.0137
新元古代氧气含量上升的假说(即新元古代氧化事件)一直被反复地与动物生命的起源和兴起联系在一起。然而,在过去十年中出现了一批新的工作,对该假说提出质疑。本文探索了现有大气和海洋氧化的代用资料,并考虑到每个地球化学系统的独特系统分类学,试图使数据相容。本文还提出了一个综合性的 COPSE 生物地球化学模型的新结果,该模型结合了几个最新的研究,创建了一个850-250 Ma 的连续模型记录。本文的结论是,新元古代氧气水平介于埃迪卡拉纪和早古生代之间,具有高度动态性,直到晚古生代才达到稳定的现代状态。因此,本文认为新元古代氧化窗口和古生代氧化事件是地球大气和海洋中氧气上升的更合适的描述。
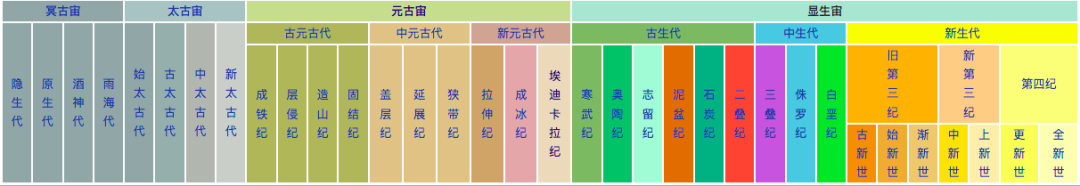
图2. 地质年代 | 来源:维基百科
2.论模型在认识复杂生命起源中的应用
论文题目:On the use of models in understanding the rise of complex life
作者:Timothy M. Lenton
发表时间:2020年6月12日
论文链接:https://royalsocietypublishing.org/doi/10.1098/rsfs.2020.0018
最近,人们提出了几个看似不相容的模型,用以描述地球系统过程与复杂生命兴起之间的关系。这些模型对元古代大气氧气水平和海洋营养水平,它们是否限制了复杂生命,以及复杂生命的兴起如何影响生物地理化学条件提出了不同的看法。对于模型应用者来说,很难评估哪个模型及其结果更加可信,因此本文对此进行了研究。本文简要回顾了相关的假说,模型如何被用来体现或检验这些假说,以及生物地球化学循环模型应该体现的关键原则。然后本文批判性地回顾了生物地球化学模型在以下方面的应用:从代用资料推断关键变量;重建古代生物地球化学循环;以及研究复杂生命如何影响生物地球化学循环。已发表的模型结果表明,元古代大气中的氧含量和海洋中的磷水平长期稳定在极低的水平。本文解释了这些问题的根源,并强调了需要解决的关键实证不确定性。最后,本文给出了如何将模型和数据更好结合的建议,以促进对地球系统过程和复杂生命兴起之间关系的科学理解。
3.早期动物进化中的生存和选择偏差以及分子钟系统性高估的一个来源
论文题目:Survival and selection biases in early animal evolution and a source of systematic overestimation in molecular clocks
作者:Graham E. Budd,Richard P. Mann
发表时间:2020年6月12日
链接:https://royalsocietypublishing.org/doi/10.1098/rsfs.2019.0110
许多假说试图解释一些如寒武纪大爆发之类的重要进化事件:为什么它们会在那个时候发生?是什么塑造了这些进化事件,为什么它们最终会走到尽头?然而,很少有人注意到“零假设”的概念,即这种多样化的某些特征仅仅是通过它们的统计结构产生的。这些统计特征似乎也影响了我们对这些事件发生时间的感知。在这里,本文特别指出,对异常大的演化支的研究,导致系统性高估了某些分子钟得出的演化支年龄,这种影响的大小可能足以解释这些分子钟和化石记录之间令人费解的错误匹配。本文对埃迪卡拉纪晚期至寒武纪的化石记录的分析表明,这段时期很可能记录了两侧对称动物真正的进化辐射,而涉及两侧对称动物各种各样神秘起源的解释似乎没有必要。
4.从化石记录看真核生物
论文题目:Insights into eukaryogenesis from the fossil record
作者:Susannah M. Porter
发表时间:2020年6月12日
链接:https://royalsocietypublishing.org/doi/10.1098/rsfs.2019.0105
长期以来,真核生物起源(Eukaryogenesis),即真核细胞的形成过程一直困扰着科学家。人们一直认为化石记录对说明这一过程几乎没有贡献,一部分原因是细胞核和线粒体等重要特征很少保存下来,另一部分原因是早期真核生物的主流模型暗示,真核生物起源发生在化石记录中确认的第一批真核生物出现之前。在这里,本文对早期真核生物进化提出了一个与主流所不同的假说。本文认为,冠群真核生物并非起源于古元古代晚期,在原核生物主导的世界里作为生态上的次要成分长达5亿多年时间,而是起源于中元古代晚期,这意味着可以通过化石记录来研究真核生物起源。本文回顾了四个冠群特征的代用资料:脱囊结构的存在证明了形成包囊的能力;棘刺结构指示了复杂的细胞骨架的出现;甾烷的存在表明了甾醇合成过程的实现;线粒体的出现及真核生物生活在有氧环境证明了有氧呼吸的发生。另外,这些特征的演化顺序也可以利用代用资料来推断。记录表明,包囊形成和复杂的细胞骨架出现在古元古代晚期,甾醇合成出现在中元古代晚期或新元古代早期。有氧呼吸的起源不容易确定,但目前的证据表明,它有可能是在中元古代的某个时候进化的。
5.吞噬作用在地球历史上的起源
论文题目:The origin of phagocytosis in Earth history
作者:Daniel B. Mills
发表时间:2020年6月12日
链接:https://royalsocietypublishing.org/doi/10.1098/rsfs.2020.0019
吞噬作用,或“细胞吞食”,是一种真核生物特有的过程,其中颗粒物被内陷的细胞质膜吞没。几十年来,吞噬作用的起源一直位于真核生物起源的讨论中心,有人认为吞噬作用是获得线粒体的先决条件或后果。然而最近,基因组学和细胞学证据越来越多地支持另外一种观点,即前线粒体宿主细胞(“Asgard”古细菌超门中的一个真正的古细菌演化支)不能吞噬细胞,而是使用替代机制来吸收线粒体的变形杆菌祖先。事实上,真核生物树上与吞噬体相关的蛋白质的多样性和变异性表明,吞噬作用可能在真核冠群中独立进化了好几次,正如在各种现存的真核生物中看到的那样。由于吞噬作用对于现代海洋食物网的运作至关重要(没有它,就不会有微生物环或动物生命) ,吞噬作用的多个晚期起源可以帮助解释为什么新元古代的许多生态和进化创新(例如,真核生物矿化的出现,“藻类的崛起”,和动物的起源)在那时出现。
6.埃迪卡拉纪早期瓮安类胚胎化石中的细胞核保存、细胞核的实验化石形成学及对解释真核化石记录的启示
论文题目:Nucleus preservation in early Ediacaran Weng’an embryo-like fossils, experimental taphonomy of nuclei and implications for reading the eukaryote fossil record
作者:Weichen Sun,Zongjun Yin,John A. Cunningham,Pengju Liu,Maoyan Zhu ,Philip C. J. Donoghue
发表时间:2020年6月12日
链接:https://royalsocietypublishing.org/doi/10.1098/rsfs.2020.0015
鉴定细胞器化石的挑战长期以来一直阻碍着解释早期真核生物进化的化石记录。本文通过现存真核生物细胞核的化石形成实验和对早期埃迪卡拉纪瓮安生物群胚胎样化石中假象细胞核的微观物理和化学特征的表征来探索这一挑战。细胞核化石具有不同的保存方式,在形状、是否存在内部矿体以及伴生矿化的化学性质等方面都有所不同。这些细胞核并不直接形成化石,而是以外部模具(mould)的形式出现。洋葱(Allium cepa)表皮细胞的化石形成实验表明,细胞核比宿主细胞具有更强的抗衰变能力,一般能够在死后数周至数月内保持其物理尺寸,尽管在某些实验条件下,细胞核会萎缩,或被微生物膜所覆盖。化石和实验证据可以在细胞质选择性矿化的单一化石形成途径中合理化,保留既能抵抗衰变又能抵抗矿物质复制细胞核的外部霉菌。综上所述,本文的结果既提供了瓮安细胞核的可靠鉴定,也提供了细胞器化石记录的可能性,这可能有助于在长期存在的关于真核级细胞进化组合的相对和绝对时间的争论中作出选择。
7.铝硅酸盐包层保存了大约8亿年前的复杂生命
论文论文题目:Aluminosilicate haloes preserve complex life approximately 800 million years ago
作者:Ross P. Anderson,Nicholas J. Tosca,Gianfelice Cinque,Mark D. Frogley,Ioannis Lekkas,Austin Akey,Gareth M. Hughes,Kristin D. Bergmann,Andrew H. Knoll,Derek E. G. Briggs
发表时间:2020年6月12日
链接:https://royalsocietypublishing.org/doi/10.1098/rsfs.2020.0011
产于泥岩中的微化石是元古代化石记录的主要组成部分,并且特别记录了早期真核生命。早期生物没有生物矿化部分来抵抗衰变,而且对于它们在泥岩中的石化过程的控制也知之甚少。因此,元古代化石记录受到损害ーー我们不知道微化石出现的时空格局的变化是否反映了进化或有利化石条件的分布。我们调查了 Lakhanda 群(俄罗斯)1000 Ma内的石化,以及 Svanbergfjellet 和 Wynniatt(斯瓦尔巴德群岛和加拿大北极圈)大约800Ma的地层。采用聚焦离子束铣削技术从薄片上提取微化石及其周围基质的垂直切片。元素面扫描和基于同步加速器的红外显微光谱显示,微化石被富含铝(可能存在于高岭石中)的包层(haloes)所包围。高岭石已经牵涉到寒武纪 Burgess 页岩类型(BST)的石化,并已知可以减缓分解者生长。新元古代泥岩微化石记录可能偏向于有利于高岭石形成的热带环境。但这些沉积物缺乏后生动物化石,尽管它们与能够保存非矿化后生动物的更年轻的 Burgess 页岩类型沉积物具有同样的石化条件。因此,后生动物,至少是那些通常保存在 Burgess 页岩类型沉积物中的后生动物,在大约800 Ma 之前可能并不存在于沉积环境中。
8.内华达州埃迪卡拉纪森林峡谷地层中富含粘土的硅塑性沉积物中 erniettomorph 化石的保存
论文题目:Preservation of erniettomorph fossils in clay-rich siliciclastic deposits from the Ediacaran Wood Canyon Formation, Nevada
作者:J. G. Hall,E. F. Smith,N. Tamura,S. C. Fakra,T. Bosak
发表时间:2020年6月12日
链接:https://royalsocietypublishing.org/doi/10.1098/rsfs.2020.0012
保存完好的三维埃迪卡拉化石出现在全球的砂岩层中。内华达州蒙哥马利山脉埃迪卡拉纪森林峡谷地层(WCF)的砂硅质碎屑沉积物,包含两种形状和大小相似的化石形态:一种具有毫米级的隆起和明显的下界,另一种则没有这些特征。本文把它们解释为 erniettomorph 的带状同形体(tapho morphs),这是一种软体生物,在分类学上具有不确定的亲缘关系。通过岩相学、拉曼光谱、 X射线荧光光谱仪微探针和 X 射线衍射等方法,对这两种非同形态的铸模(cast-and-mould)保存进行了研究。所有化石及其周围的沉积基质都含有石英颗粒、富铁绿泥石和白云母。脊状化石中的石英颗粒比无隆起的同形体多大约70% ,表明脊状化石中粘土矿物丰度较低。绿泥石和白云母可能起源于蒙脱石和高岭石前驱体(precursor),经历了绿色片岩相变质作用。据推测,脊状化石周围的沉积物中含有丰富的高岭石和蒙脱石,这使得脊状化石保存了一个连续的、明显的、富含粘土和油母质的底界。在森林峡谷地层隆起的化石和其他地点的 erniettomorph 中发现的石英,也表明这种矿物在砂岩和粉砂岩矿床的 erniettomorph 的三维保存中发挥了作用。
9. 动物的起源和早期进化:从遗迹化石的角度看我们处于什么位置?
论文题目:The rise and early evolution of animals:where do we stand from a trace-fossil perspective?
作者:M. Gabriela Mángano,Luis A. Buatois
发表时间:2020年6月12日
链接:https://royalsocietypublishing.org/doi/10.1098/rsfs.2019.0103
遗迹化石记录为追踪动物的起源和早期进化提供了丰富的信息。它包含了硬体和软体生物的活动,在埃迪卡拉纪(635-539 Ma)-寒武纪(539-485 Ma)的过渡期间是连续的,包含动物行为和它们作为生态系统工程师的角色等深刻信息,并且允许对古环境背景有更精确的特征刻画。为了揭示遗迹化石记录中的宏观进化信号,可以采用多种方法,包括生物扰动程度的估计,遗迹多样性,遗迹差异轨迹分析,以及对动物生态空间占有情况和生态系统工程类型的评价。
对遗迹化石记录的分析表明,在埃迪卡拉纪存在能动的底栖两侧对称动物,主要以生物膜为食。尽管埃迪卡拉纪遗迹化石简单且位于沉积物表面或表面下方,但在埃迪卡拉纪末期,遗迹化石的复杂性、捕食压力、沉积物扰动和穿透深度的增加是显而易见的。尽管如此,在寒武纪早期,遗迹化石的多样性和差异性显著增加,这强调了幸运期(Fortunian,539-529 Ma,埃迪卡拉纪之后的寒武纪的第一个阶段)的新颖性不可低估。幸运期仍然显示了埃迪卡拉纪式的基地生态的持久性,但其根本特征是新的遗迹化石建筑计划,反映了与基质相互作用的新方式。显生宙式底栖生态系统的出现证明,食物网的长度和连通性增加,有机碳转移和养分循环的效率提高。在寒武纪早期,无论是高能砂质近岸环境还是低能泥质近岸环境,动物内部生态空间都发生了深刻的重组。
本文提出了一个包括四个演化阶段的模型,综合了埃迪卡拉纪-寒武纪的遗迹化石记录的信息。研究人员使用丰富的遗迹学工具,对埃迪卡拉纪-寒武纪遗迹化石记录进行了严格、系统和全面的评价,对遗迹学数据集和沉积学信息进行了高分辨率的整合,表明生源混合的出现是显生宙初期完全海洋环境的一个重要因素。
10.埃迪卡拉纪的生物层积学:埃迪卡拉纪生物的沉积环境、生态学和生物学意义
论文题目:Biostratinomy of the Ediacara Member (Rawnsley Quartzite, South Australia):implications for depositional environments, ecology and biology of Ediacara organisms
作者:Mary L. Droser,Lidya G. Tarhan,Scott D. Evans,Rachel L. Surprenant,James G. Gehling
发表时间:2020年6月12日
链接:https://royalsocietypublishing.org/doi/10.1098/rsfs.2019.0100
前寒武纪埃迪卡拉纪生物群ーー地球上最早的宏观多细胞生物群落的化石记录ーー为我们这个星球上复杂生命的出现提供了重要见解。在南澳大利亚的埃迪卡拉化石遗址Nilpena发掘和重建了 Rawnsley Quartzite 中的埃迪卡拉系近300平方米的包含化石的层理面,使人们能够对埃迪卡拉化石组合的沉积学、化石形成学和古生态学进行详细研究。埃迪卡拉大型化石和具有纹理的有机表面在岩相、层面和个别标本的特征刻画,已经对埃迪卡拉纪群落所居住的古环境(特别是水动力条件)产生了前所未有的洞察力,影响了埃迪卡拉生物的自体和共生态,以及埃迪卡拉化石的形态和组合组成。
本文描述了环境过程如何介导了埃迪卡拉化石组合的沉积相的发育。以 Arborea 和 Dickinsonia 这两种最常见的埃迪卡拉系化石为例,本文勾勒出了可用于区分生态、环境和生物结构信号的标准,并重构了这些过程之间的相互作用如何明显地塑造了埃迪卡拉化石记录。
11.环境条件对埃迪卡拉纪生物群落交错区的影响
论文题目:The influence of environmental setting on the community ecology of Ediacaran organisms
作者:Emily G. Mitchell,Nikolai Bobkov,Natalia Bykova,Alavya Dhungana,Anton V. Kolesnikov,Ian R. P. Hogarth,Alexander G. Liu,Tom M. R. Mustill,Nikita Sozonov,Vladimir I. Rogov,Shuhai Xiao,Dmitriy V. Grazhdankin
发表时间:2020年6月12日
链接:https://royalsocietypublishing.org/doi/10.1098/rsfs.2019.0109
大尺度环境对现代海洋生态系统的形成起着重要作用,但古生物群落受其环境影响的程度尚不清楚。为了研究大尺度环境对早期动物生态系统群落交错区的影响,本文采用空间点过程分析(spatial point process analyses,SPPA)方法,对来自不同环境和不同地点的7个埃迪卡拉纪晚期(558-550Ma)的层面组合进行了研究。所研究的古生物群落对海底亚米尺度生境异质性的反应表现出明显差异:浅海(近岸)古生物群落受到局部生境异质性影响严重,而深海古生物群落则不同。浅水群落中的局部差异可能因食草动物和腐生动物的存在而进一步加剧,这些动物的行为可能导致底栖群落从浅海到深海沉积环境的生境异质性日益增加。在所研究的时间间隔内,埃迪卡拉纪浅水群落的物种丰富度高于深水群落,因为生境的异质性增加了现代海洋环境中的物种丰富度。本文的研究结果为早期动物多样化的“热带稀树草原(Savannah)”假说提供了定量支持,即埃迪卡拉纪的生物多样化由局部底栖环境的空间差异驱动。
12.利用缺氧条件作为复杂多细胞生物进化的驱动力
论文题目:Harnessing hypoxia as an evolutionary driver of complex multicellularity
作者:Emma U. Hammarlund
发表时间:2020年6月12日
链接:https://royalsocietypublishing.org/doi/10.1098/rsfs.2019.0101
细胞组织的维持需要低氧条件。对低氧环境的需求,与细胞分化随着氧气环境的扩大而出现进化飞跃的想法形成了鲜明对比。然而,脊椎动物和维管植物为了适应氧气条件下的组织更新,表现出了适应缺氧的能力。本文提出大型(多样化)生命需要缺氧条件的假设,并给出了相应证据及该假说的影响。利用缺氧条件的能力使得多细胞生物进化至能够离开氧气浓度稳定的小生境,去往氧气浓度波动的新环境。由于在地球表面的大多数环境中氧气浓度都是波动的,多细胞最初的小生境可能是在深海环境中。该假设解释了大型(多样化)生命利用缺氧条件促进干细胞更新、维持组织,并解释了多细胞生物在地球历史上大部分时候稀有的原因。
13. 风化、变质和重建地球的氧化作用
论文题目:Weathering, alteration and reconstructing Earth’s oxygenation
作者:Noah J. Planavsky,Leslie J. Robbins,Balz S. Kamber,Ronny Schoenberg
发表时间:2020年6月12日
链接:https://royalsocietypublishing.org/doi/10.1098/rsfs.2019.0140
氧气含量在控制复杂生命进化辐射的时间和速度方面起到了怎样的作用(如果有的话) ,回答这个问题是地球科学和生命科学中最基本的问题之一。准确地重建地球的氧化还原历史是解决这个问题的一个重要部分。在过去几十年里,大量研究利用地球化学氧化还原的代用资料来重新讲述地球氧化作用的故事。
然而,其中许多研究,甚至那些采用相同地球化学代用资料的研究,对氧化事件的时间和强度产生了矛盾的解释。对于相互冲突的氧化还原重建,有两种可能的解释:(i)氧气含量在时间和空间上都具有令人难以置信的动态性;(ii)包括本文作者在内的研究人员群体,我们经常研究受次生风化和变质(特别是次生氧化)影响的岩石,却忽略了这种改变对生成数据的影响。
现在有多个案例研究记录了以前忽视的次生变化,解决了一些与氧化还原进化冲突的约束。在这里,对一个页岩地球化学大型数据库的分析,揭示了岩石样品和钻孔岩心样品之间铈(Ce)异常的显著差异,一种常见的古氧化还原代用资料。这种不一致性支持了这样一种观点,即受次生作用影响的样品的地球化学数据经常发表在同行评审的文献中。
无论是个人还是整个地球化学界,我们大多数人都没有及时意识到这个问题有多么普遍,但其他社群也面临过此类质量控制危机。进一步的证据还有,深时地球化学样品极易受次生作用影响,并且这种次生作用可能导致虚假结果和对古环境的虚假解释,这表明需要将样品数据在公开访问的数据集中存档作为发表新的古氧化还原数据的先决条件。最后,地球化学界需要思考如何实施更多的质量控制措施,以提高古氧化还原代用资料的准确性。
14.复杂生命的进化与地球系统的稳定
论文题目:The evolution of complex life and the stabilization of the Earth system
作者:Jonathan L. Payne,Aviv Bachan,Noel A. Heim,Pincelli M. Hull,Matthew L. Knope
发表时间:2020年6月12日
链接:https://royalsocietypublishing.org/doi/10.1098/rsfs.2019.0106
在5亿年动物进化的历史中,物种的背景灭绝率在降低。地球逐渐变得适宜动物生存的原因可能源于以下几个过程:(i)导致物种灭绝的物种间相互作用的强度下降;(ii)洪水、火山喷发等地质灾害的发生频率或强度下降;(iii)动物对环境干扰的敏感度下降;或(iv)气候系统和生物地球化学循环中稳定反馈的强度上升。
没有证据表明,随着时间的推移,物种之间的相互作用或地质灾害发生频率或强度有所下降。然而,来自古生物学、地球化学和比较生理学的证据表明,动物已经变得更能适应环境的变化;而且总的来说,复杂生命的进化加强了气候系统中稳定的反馈。
某些门类非比寻常的成功似乎至少部分是源于埃迪卡拉纪和寒武纪时期,要进化到宏观尺寸的解剖学上的解决方案。身体尺寸较大的动物,由于解剖复杂性的增加,越来越能够混合海洋沉积物和水柱,从而促进生物地球化学循环的稳定性。此外,促进生态分化的身体架构往往与较低的灭绝率有关。从这个意义上说,寒武纪问题的解决方案对复杂生命的轨迹产生了持久影响,反过来,也影响了地球系统的基本属性。
复杂科学最新论文
集智斑图顶刊论文速递栏目上线以来,持续收录来自Nature、Science等顶刊的最新论文,追踪复杂系统、网络科学、计算社会科学等领域的前沿进展。现在正式推出订阅功能,每周通过微信服务号「集智斑图」推送论文信息。扫描下方二维码即可一键订阅:

推荐阅读
-
生命形式曾经多次发生?多重视角探寻生命起源的理论结构 -
生命起源百年探索:进化论、信息论、热力学谁是主角? -
热力学怎样理解生命:熵、信息、自由能和复杂性的模糊图景 -
生命如何在复杂环境中生存?信息、调控和几何结构的交织 -
什么是生命起源 Abiogenesis | 集智百科 -
《张江·复杂科学前沿27讲》完整上线! -
加入集智,一起复杂!
点击“阅读原文”,追踪复杂科学顶刊论文
微信扫一扫,分享到朋友圈











