今天的地球处于人类世,人类活动对整个地球生态系统具有深刻影响。由于干旱和过度放牧,草原转变为沙漠;由于滥砍滥伐和火灾,热带雨林转变为稀树草原——一旦外部扰动超过限度,生态系统就会越过临界点,从一个平衡态转变到另一个相对较次的平衡态,难以恢复原本的一派生机。人类是否可以找到早期预警信号,规避系统临界点的到来?
空间斑图*形成(Spatial pattern formation)被认为是包括生态系统在内的复杂系统中危险的临界点(tipping point)和即将发生的临界转变(critical transition)的早期预警信号。Rietkerk等人2021年10月发表于 Science 的综述文章《通过空间斑图形成避免复杂系统崩溃》综述了生态系统和地球系统的组分在实际中如何通过多种空间斑图形成的方式规避灾难性的临界点。利用数学方法和实际案例,他们认为,许多具有临界倾向的生态系统和地球系统组分都可能出现规避临界点和增强恢复力的现象。由于过去被忽视的空间动力学和多种稳定状态的存在,许多复杂系统可能比原先认为的更具恢复力,因此可能不会随着气候变化发生灾难性的转变。
*译者注:本文将 spatial pattern 与 Turing pattern 中的 “pattern” 一词统一翻译为“斑图”,在生态学领域中,pattern 也常翻译为“格局”。
研究领域:空间生态学,临界点,图灵斑图,空间自组织
Max Rietkerk et al. | 作者
吕丽莎、胡一冰、李明章、郭瑞东、张澳 | 译者
张澳、梁金 | 审校
邓一雪 | 编辑
论文题目:
Evasion of tipping in complex systems through spatial pattern formation
https://www.science.org/doi/10.1126/science.abj0359
目录
结构化摘要
摘要
一、生态系统临界现象及其预警信号
二、通过图灵斑图规避临界点
三、通过共存态规避临界点
四、走向空间斑图形成及多稳态的理论
五、典型系统:规避稀树草原的临界点
六、空间斑图形成及多稳态的普遍条件
结论
在人类世中,我们需要更好地理解气候和土地利用的变化对生态系统、地球系统组分和全部地球系统的灾难性影响。临界转变的概念,即从一个状态向另一个状态的转换,有助于这一理解。临界转变意味着系统被迫离开原有平衡态的吸引域,进入另一种通常较次的稳定状态。在这一情况下,对于此类即将发生的临界转变的早期预警信号的搜寻已经成为研究的关注点。特别是,生态系统中的空间自组织,比如规则的植被斑图,即图灵斑图(Turing pattern)的自发形成过程被认为是一个重要的早期预警信号。
然而,最近的发现指出,这种空间自组织未必应该被解释为临界转变的早期信号。反之,空间自组织可以使生态系统规避临界点,因此可以成为其具备恢复力的信号。这些发现源于最近对空间模型的数学分析和对现实生态系统的观察。它们都揭示了多稳定性(multistability),意味着许多不同的空间斑图可以在同一环境条件下共存,并且每个斑图都可以在较大范围的条件内保持稳定。这使得复杂系统的状态在临界点之外仍可以通过空间自组织而维持稳定。此外,如果一个具有临界属性的复杂系统经历了扰动,后续的变化未必会使整个系统发生临界转变。反之,这种变化可以保持在局部范围,因为生态系统允许多个状态共存于空间中,这被称为共存态(coexistence states)。通过这种途径,即使环境恶化,空间状态也可以维持于临界点以外。我们将图灵斑图和共存态称为空间斑图形成。许多原先被认为接近临界状态的生态系统和地球系统组分(包括整个地球系统),可能通过多种空间斑图形成过程规避了临界点。
为了进一步研究复杂系统如何通过空间斑图形成来规避临界点,稀树草原系统因其被观测到的可选状态和空间斑图,可以用来作为一个典型案例开展具体研究。另外,在生态系统和地球系统组分中,都可以用数学方法分析规避临界点的普遍条件。通过研究何种小幅度的图灵斑图可以增长成为大型的局部交互结构,可以找到帮助复杂系统规避临界点的相关场景。我们应当研究全球变化对于共存态间的空间边界的影响,并且揭示空间域的限制和人类带来的局部和非局部的同质化效应。这一方法将会增加我们对自然中临界转变的理解和预测能力,并且能够揭示其中哪些可以被规避乃至反转。
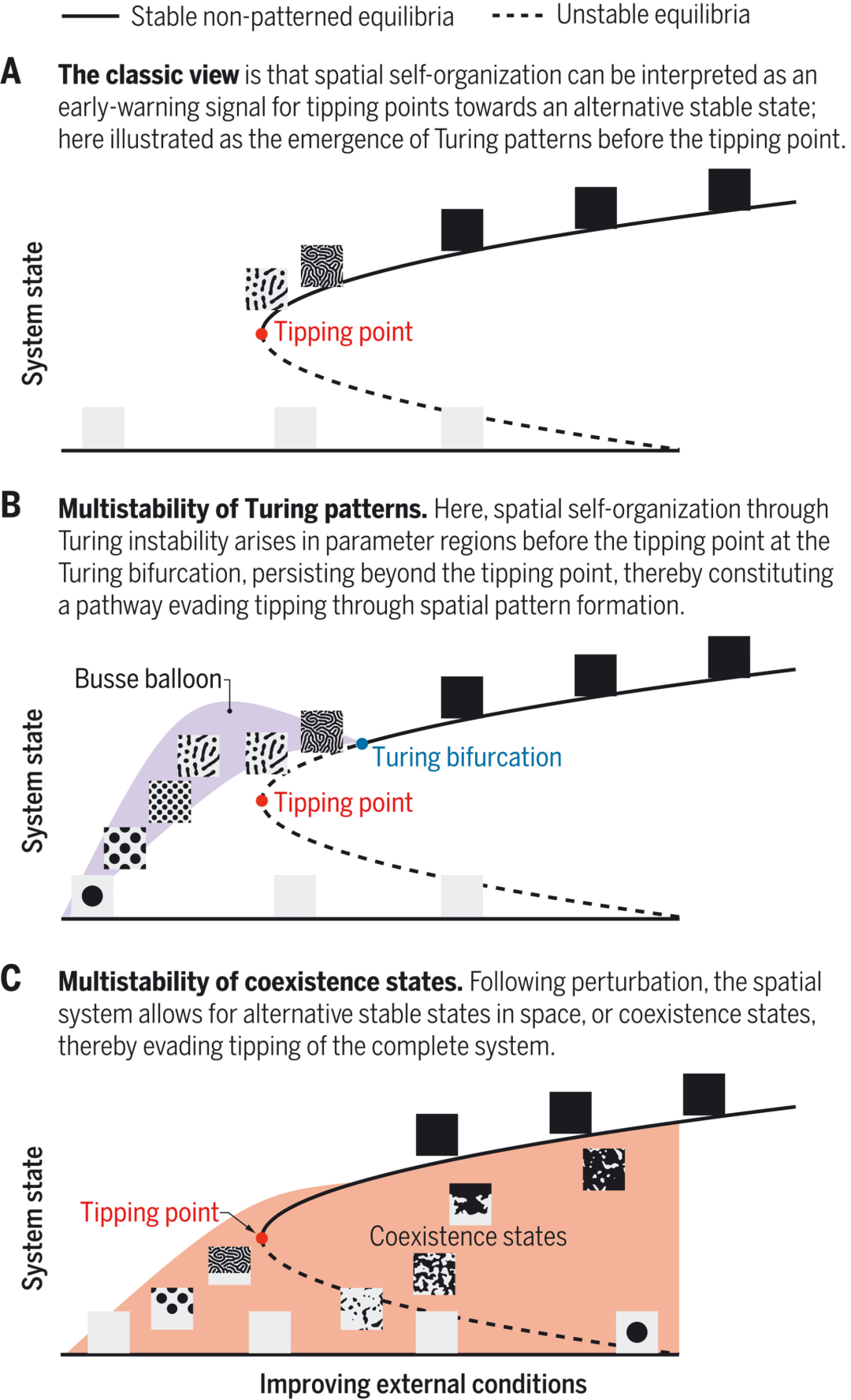
图0. 临界点的规避。我们描绘了复杂系统对外部条件的响应(即分岔图)。灰黑色方块代表了系统稳定变量的高密度状态,亮灰色方块代表了其低密度状态。(A)经典视图,(B)图灵斑图的多稳定性。借助卫星对实际的生态系统的观测,最近的模型分析揭示了图灵斑图在Busse ballon中的多稳定性。Busse ballon 是数学模型的参数空间中斑图平衡(patterned equilibria)的多稳定性出现的区域。(C)共存系统的多稳定性。规避临界点也可以是共存态的多稳定性的结果。这些空间斑图最早产生于临界点前的双稳定性区域;在外部条件恶化时,演化的空间斑图也可以维持于临界点以外,从而构成另一种规避临界点的方式。
A. 经典视图:空间自组织可以被认为是一种从临界点转向另一种稳定状态的早期预警信号;这里描绘了临界点出现之前图灵斑图的出现。
B. 图灵斑图的多稳定性。这里,在图灵分岔图的临界点之前,空间自组织通过图灵不稳定性出现于参数区域中,在临界点之外仍然保持稳定,从而通过空间斑图形成的方式规避临界点。
C. 共存稳定态的多稳定性。在扰动后,系统允许空间中存在可选稳态,或共存态,从而规避了整体系统的转变。
临界点和临界转变的概念可以帮助我们理解全球变化可能对我们的生态系统、地球系统组分(包括整个地球系统)带来的灾难性影响。研究者们正在寻找早期的预警指标,其中空间自组织就是一个信号。本文综述了空间自组织如何帮助复杂系统规避临界点,从而成为恢复力的信号。许多至今仍被认为具有临界倾向的生态系统和地球系统组分(包括整个地球系统)可能通过不同的空间斑图形成过程来规避临界点。我们提出了一种系统分析,可能会揭示规避临界点和形成恢复力的广泛条件范围。
临界转变和临界点的概念有助于我们理解人类世的地球变化[1-3]。这一观点认为,经历全球变化的生态系统或地球系统组分通常能够持续保持平衡状态。然而,这都只发生于达到临界点之前。在临界点,期望的稳定态会消失,系统会经历一个临界的灾难性转变,达到一个新的平衡态[1,2,4]。后者所处的状态常伴随外部环境恢复到初始值,并展现出迟滞现象(hysteresis)[1,5]。比如说生态系统中,由于营养物质过多导致清澈的湖泊变得浑浊[6];由于干旱或过度放牧,贫瘠的沙漠取代了干旱的稀树草原或旱地的植被区域[5,7];由于滥砍滥伐、火灾或降水量减少,热带稀树草原取代热带森林[8-10]。
导致这种转变的动力学机制是正反馈(positive feedbacks),正反馈过程会放大施加在复杂系统上的变化。在上述生态系统的案例中,正反馈使湖泊的浑浊度增加,导致大型植物减少、养分吸收被抑制,湖水从而变得更加浑浊[6];干旱稀树草原的植被覆盖减少,导致土壤水分渗透减少,进一步加剧植被消失[5,7];滥砍滥伐和火灾导致森林树木减少,杂草肆意生长引发更多的火灾,从而导致森林树木进一步减少[10-12]。在临界点时,正强化反馈(positive reinforcing feedbacks)压倒了维持生态系统理想状态的负平衡反馈(negative balancing feedback)过程,这往往导致生态系统功能的灾难性损失。
有许多可能的途径可能让正反馈超过负反馈并触发状态转变。这种转变是生态系统恢复力退化的结果,恢复力可以被理解为一个系统状态在发展到临界点之前所能承受的环境变化量或扰动强度[13]。三种最可能的临界情况是[14]:
1)分岔导致的临界(Bifurcation-induced tipping,B-临界)出现是因为,一个参数的移动(如环境变化)使得初始稳定状态的吸引域(basin of attraction)降低为零;
2)噪声导致的临界(noise-induced tipping,N-临界)出现是因为,系统状态的扰动(如环境噪声或干扰)将系统推到初始状态的吸引域之外;
3)变化速率导致的临界(rate-induced tipping,R-临界)出现是因为,环境参数的变化快于恢复至初始状态的速度。
在生态学文献中,系统恢复到初始状态的吸引力,或恢复平衡状态的速度,即为系统的恢复力或稳定性[13]。
这种临界转变的思想不仅对局部尺度的生态系统很重要,对于许多区域尺度的地球系统组分也同样重要[15,16]。比如:气候变暖引发的北极冰盖消融会导致冰面反射率减小,从而加剧气候变暖[17-19];地表水通量的变化导致大西洋环流变化[20-22]。从生态系统到气候变化,临界点这一概念可应用到多种尺度。例如在全球尺度上,临界点的概念可以用于定义发生全球性灾难事件的环境边界条件[23],同样,也可以用于研究整个地球生物圈对气候和土地利用变化[3,24]的响应。
对于像旱地、稀树草原和泥炭地这些在空间延伸的生态系统,已有研究强调了关键的状态转变与植被自组织空间斑图的形成有关[2,25]。随着环境条件的恶化,这些系统中原本覆盖均匀的植被变得不再均匀,从而形成一些有规则的空间斑图。这种均匀状态的空间不稳定性被称为图灵不稳定性(Turing instability)或图灵分岔(Turing bifurcation),最早由计算机和人工智能之父艾伦·图灵在研究反应-扩散系统时发现并命名[26]。在图灵不稳定性观点诞生之后,所谓的图灵斑图(Turing pattern)也应运而生,即在不同的环境条件下,会产生不同的空间斑图。在之前的研究中,图灵斑图被认为是临界点走向生态系统另一状态的助推[25]。按水分、食草情况等环境条件逐渐恶化的顺序排列,常见的(尤其在旱地中)植被斑图有以下几种:存在裸露间隔的均匀植被、迷宫般或条状的植被、带有点状植被的裸露土地[25,27-30]。
图灵不稳定性的机理是前文提到的正反馈,它在空间延伸的系统上具有尺度依赖性:正反馈逐渐减弱,并在空间中更远的地方被负反馈逐步取代,在这个空间延伸过程中便产生了尺度依赖的反馈作用。而当假设系统同质时,这些导致尺度依赖反馈的过程的关键空间效应通常被忽略[31]。例如在旱地,尺度依赖的反馈作用涉及到水下渗量及土壤吸水量,在植被生长区这两者更高。这导致植被会吸取更远处的可用水,让地表水和土壤水流向植被,从而形成某处植被密集,某处土壤贫瘠的空间斑图。
到目前为止,图灵不稳定性导致的规则空间斑图被认为是各种生态系统中临界点或向可选状态转变(B-临界)的早期预警信号[2,25]。然而,最近结合卫星观察和数理分析的生态系统研究提出了一种新观点[32-34],揭示了 Busse balloon (根据F. H. Busse[32,33,35]命名)理论阐述的多稳定性的存在。Busse balloon(如图B紫色区域)对应模型中参数空间的一个区域,对于该区域,大范围波长的规则空间斑图(干旱草原示例中的植被)可能存在且保持稳定。此外,当模型中的条件或参数发生变化时,特定的空间组织可以保持稳定,这意味着在到达 Busse balloon 边缘之前,波长在各种环境条件下都不会改变。空间组织和生态系统状态变量(如生态系统生产力)可以通过非临界或非灾难性的方式微调(如图1B)。在非空间模型中,均匀植被状态可能仍然存在,或者已经退化为裸露土壤。
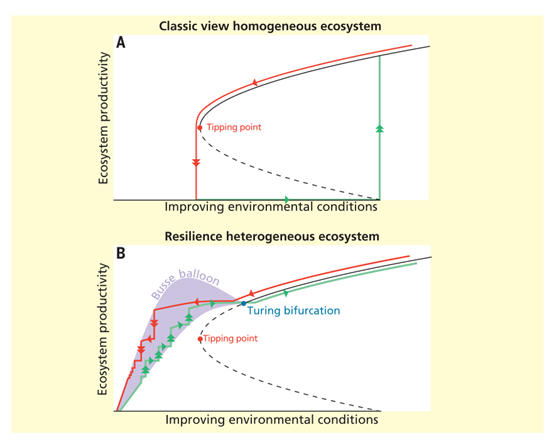
图1. 同质和异质生态系统的恢复力。实线表示稳定平衡,虚线表示不稳定平衡。双箭头表示生态系统状态转变,单箭头表示生态系统状态微调。生态系统功能退化(红色)和恢复过程(绿色)的典型轨迹,显示了具有异质和空间自组织特征的生态系统的迟滞回路。
(A)同质生态系统中的恢复力。通常认为,不断恶化的环境条件会造成生态系统状态或生态系统生产力的微小调整,直到系统状态越过临界点(红点)后发生关键或灾难性的转变。
(B)异质生态系统中的恢复力。在多稳态空间生态系统中,图灵分岔点(蓝点)之外的任一环境条件,都会驱动系统进入多重稳定、空间自组织状态,即图中的紫色阴影区域(也称为Busse balloon)。此种情形下,生态系统中会发生很多的小转变,而非临界转变。这些小转变对生态系统整体的功能和生产力的影响比较小。
上述的新观点源于这样一个事实,早期对生态系统模型的分析忽视了空间斑图状态的许多可能响应和Busse balloon的存在。而这一超越传统的研究考虑了斑图状态的稳定性[32],体现了Busse balloon思想,从而有了更贴切真实生态系统的新发现。
最近,这些包含多稳态、Busse balloon 的模型预测在真实生态系统观测中得以印证[33]。根据卫星观测显示,在非洲干旱大草原不同地域范围内均出现有规律的空间植被斑图。这些观测结果表明,在环境条件相似的同一区域内,以及在同一较大范围内的不同子区域内,许多不同波长的空间斑图彼此相邻,这一现象也支持了 Busse balloon 的存在[32,33]。此外,尽管外界环境条件发生变化,但某些特定区域的斑图波长数十年来保持稳定。我们将上述这两种现象结合起来称为“多稳态”,即在同一环境条件下,共存有多种不同的空间斑图,其中每种斑图都可以在较宽的条件变化幅度内稳定存在。此外,最近的模型分析发现在Busse balloon的边界处,生态系统会进行微调使之空间状态保留在Busse balloon内。换句话说,形成空间斑图的主导变量(例如生态系统生产力)并没有急剧突变,而是逐渐微调[34]。另外,斑图状态下的植被即便身处临界点以外也依旧能生存。因此,种种现象证实了这样一种观点:到达临界点之前,系统在图灵分岔处呈现出空间斑图,复杂生态系统的状态可以在临界点之外继续维持。
因此,曾经被认作是系统转变的早期预警信号的“图灵斑图”,现在似乎能象征系统的某种恢复能力。在这种情况下,斑图来源于图灵不稳定性,并导致斑图多稳定性和系统状态的逐渐变化。在其他类似研究中,如贻贝聚生地(见图2)和真实系统中也发现了斑图状态的多稳定性[36,37]。值得注意的是,预测的生态系统状态变化量可能不同,这取决于环境变化的幅度和速度[32,34]。
图2. 潮滩上贻贝和硅藻的空间斑图形成。这种斑图的形成能够规避海平面上升引起的临界点,这样潮汐滩就不会被淹没。其中潮汐板块的空间斑图及波浪基于数学模型模拟。| 图片来源:Johan van de Koppel/Ulco Glimmerveen
目前有明确迹象表明,通过空间自组织规避临界点可能在生态系统和地球系统组分中非常普遍。该结论基于针对真实生态系统中规则斑图形成的早期工作,本文提出的最新洞见以及Box 1 中一类模型的数学分析[31]。这也得益于其他研究的支持——临界点和可选稳定状态(双稳定性)已经在简单、非空间模型中提出,并至少适用于以下生态系统:旱地或干旱稀树草原[5,7],稀树草原[10,11,38],泥炭地[39],贝类聚生地[40],泥滩[41],堰洲岛[42]和海草生态系统[43]。最值得注意的是,对于所有这些生态系统模型,空间自组织或图灵斑图都已经在它们对应的真实生态系统中观察到,而这些系统往往都是空间延伸的[31,44-47]。
规避临界点可能不局限于Busse balloon中的图灵斑图,还包括更全面的空间斑图形成,我们将在后续介绍。除了空间斑图的形成,还有其他机制可以避免系统状态转变。例如,与状态变量的变化率相比,在相对简单的地球系统组分中快速重置变化参数可以规避临界点[48]。我们预计状态变量的空间斑图和时间延迟不仅会影响 B-临界,而且对 N-临界和 R-临界也有非常相似的影响——即规避整个系统的崩溃。在这种情况下,值得注意的是与临界点相关的阈值通常很少能从经验数据中观察到[49]。
在数学模型中,当平衡态失去吸引力并在参数的作用下消失时,就会发生B-临界,比如系统参数A超过其关键临界值A*。这时,系统不得不转向较次平衡状态[10,38,39]。在以往研究中,发生状态转变的模型通常是空间同质的,也就是非空间延伸的。然而,相关的建模系统几乎无一例外地在空间上扩展,因此很有必要对空间效应进行建模。状态转变前图灵现象的主要观点是,初始平衡态必须在状态转变前保持稳定。那么,在转变来临之前的同质平衡状态对空间效应也是稳定的吗?否则,空间延伸情况下的模型将不会出现状态转变,空间斑图出现在参数A达到其临界值A*之前。该情况下,模拟的生态系统状态将不会发生转变,而是通过形成空间斑图来规避转变。
如何确定是否发生状态转变前的图灵现象呢?我们考虑动力学过程:生物量b(t)和资源量r(t),假设其对应变化率分别为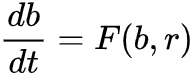 和
和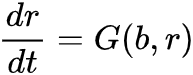 ,它们可用来反映在参数A作用下的生长、死亡和交互作用。比如说,Bastiansen关于旱地植被生长动力学的研究中[33],b(t)和r(t)分别代表植被量和水资源量,并且满足F(b,r)=-Mb+r2和G(b,r)=A-r-rb2,此处M和A表示植物死亡率和降雨量。当两个平衡态融为一体,参数A=A*时系统状态转变发生,此时系统状态为(b*, r*)。为了保证在系统状态发生转变之前平衡态是稳定的,需要满足:
,它们可用来反映在参数A作用下的生长、死亡和交互作用。比如说,Bastiansen关于旱地植被生长动力学的研究中[33],b(t)和r(t)分别代表植被量和水资源量,并且满足F(b,r)=-Mb+r2和G(b,r)=A-r-rb2,此处M和A表示植物死亡率和降雨量。当两个平衡态融为一体,参数A=A*时系统状态转变发生,此时系统状态为(b*, r*)。为了保证在系统状态发生转变之前平衡态是稳定的,需要满足: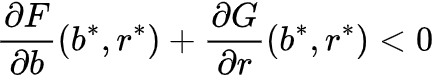 。为考虑状态转变前图灵现象是否会发生这个问题,我们需要将空间效应纳入同质化模型中。一个简单的方法是将b(t)和r(t)拓展到反应-扩散方程中,即生物量B(x,t)和资源量R(x,t)满足
。为考虑状态转变前图灵现象是否会发生这个问题,我们需要将空间效应纳入同质化模型中。一个简单的方法是将b(t)和r(t)拓展到反应-扩散方程中,即生物量B(x,t)和资源量R(x,t)满足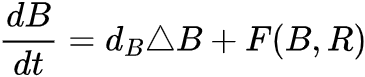 和
和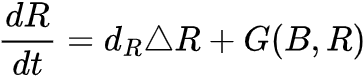 ,其中Δ是模型空间扩散量,dB和dR是控制B(x,t)和R(x,t)的扩散因子。仅可能在
,其中Δ是模型空间扩散量,dB和dR是控制B(x,t)和R(x,t)的扩散因子。仅可能在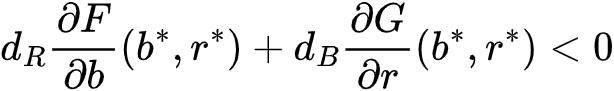 ,临界状态(b*, r*)在空间扰动下稳定得到。如果满足和
,临界状态(b*, r*)在空间扰动下稳定得到。如果满足和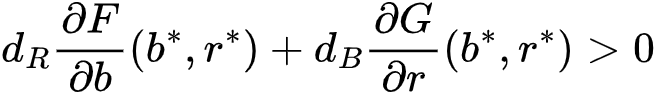 则会在状态转变前的图灵现象。因此,这个条件决定了生态系统是否可以通过形成斑图来规避临界点,并且可以在任何给定的模型中进行明确检查。比如在之前提到的旱地植被生长研究中[33],得到M<2,eM>2,此时 (b*, r*)=(1, M),A*=2 M,dR=e,dB=e。最终取值为M=0.45,e=500[33]。这样,图灵现象在转变发生前就出现了,研究的观察结果也证明了这一点。对于多变量模型和/或具有超线性扩散的空间效应模型,同样也可以推导出图灵斑图的显式条件。
则会在状态转变前的图灵现象。因此,这个条件决定了生态系统是否可以通过形成斑图来规避临界点,并且可以在任何给定的模型中进行明确检查。比如在之前提到的旱地植被生长研究中[33],得到M<2,eM>2,此时 (b*, r*)=(1, M),A*=2 M,dR=e,dB=e。最终取值为M=0.45,e=500[33]。这样,图灵现象在转变发生前就出现了,研究的观察结果也证明了这一点。对于多变量模型和/或具有超线性扩散的空间效应模型,同样也可以推导出图灵斑图的显式条件。
复杂系统可以通过空间斑图的形成来规避临界点,但这些规避临界点的条件和机制不限于导致Busse balloon 中图灵斑图形成的机制——它们还包括了更全面的空间斑图形成。虽然非空间复杂系统对扰动的响应只能是全系统范围的响应,这种响应要么导致系统完全转换到另一种稳定状态(N-临界) ,要么恢复到初始状态。但对于真正的系统来说,情况并非如此,因为真正的系统总是在空间上延伸的。在这样的系统中,无处不在的局部或随机扰动可能导致空间斑图的形成,其中只有部分空间域转换到其他状态,而全系统范围的N-临界点被规避。这样的空间斑图由空间中不同的稳定态共存而成,在此称为共存态。在给定的环境条件[50,51]下,这些共存态的不同空间斑图具有多稳定性。空间边界或表面必然会出现在空间的不同状态之间。这些边界可以是非常简单的单一空间边界,也可以是由多种稳定态组成的丰富的斑图结构。例如,冰的存在与不存在是可能发生在整个地球系统空间域中的两种不同的稳定态;这些状态在全球范围内可以共存。冰只出现在极地纬度,中间地带并没有冰,它们之间有一个简单的空间边界,称为冰线或接地线[52,53]。在较小尺度上,冰和无冰状态之间的这种空间边界可以由更复杂的可选稳定状态[54]结构构成。
划分共存态的空间边界本身可能变得不稳定,这种不稳定可能导致一个较次状态侵入另一个状态,从而提供一种机制来演化这种丰富而复杂的可选稳定态[51,55]的(空间)斑图结构。此外,在全球系统尺度上,环境变化可能导致空间斑图的重组,而系统特征不会发生突变。因此,该系统可以避免临界转变,相反[50,51],可以期待系统会有更平稳、更渐进的响应。以这种方式,空间斑图随着环境条件的恶化也可能持续到临界点后。这是另一条生态系统和地球系统组分通过空间斑图的形成和多稳定性来规避临界点的重要途径。出乎意料的是,这样的空间组织甚至可能导致非强制的反向转变,在这种转变中,系统的状态会反抗(另一种状态的)入侵,换言之,从另一种状态[55]自发地恢复至原来的状态。
这些共存态也可能在生态系统中形成,因为生物体可以聚集成自组织的斑块[56],正如在贻贝床中的贻贝集群[57]中所观察到的那样(见图2)。在这里有一个有趣的理论,与相分离(phase separation)的物理理论平行。该理论描述了相空间界面的动力学(在本文术语中,指共存态之间的空间边界),并解释了各种可能变化极缓慢的瞬态空间斑图,包括迷宫斑图、斑点斑图和粘性指进斑图(viscous fingering pattern,多孔介质中两种流体间形态不稳定的界面上形成的斑图,常见于通过土壤等介质的排水过程中,图3)。当条件改变时,这些斑图可能会导致非常平缓的过渡。导致共存态形成的相分离动力学也可能存在于生态系统和地球系统组分中[58]。例如,一些生物和资源的空间聚集可以这样解释。除了贻贝[57]外,已经对其他生态系统的相分离机制进行了比较,例如,植被在指进斑图[55]中的聚集。
图3. 两片TiO2溶胶-凝胶薄膜形成的指进斑图。| 来源:Wikipedia
目前需要发展新的理论和程序来区分空间斑图形成与现有的异质性斑图,因为真实系统通常是这两种斑图的混合物[59]。受地形控制的排水斑图是现有的异质性空间斑图的典型案例,用以解释树木的分布斑图,该例子可以在特定的植被斑图背景下伪造空间斑图形成的机制。
前文强调了复杂系统中空间斑图的形成能够规避临界点并增加系统恢复力。Box2和Box3中展示了案例。然而,某些复杂系统在形成空间斑图后,仍会呈现临界现象。这样的话,什么时候不涉及空间斑图的经典临界点理论会失效,需要新的基于空间斑图的理论作为补充?对于特定生态系统及地球系统中的组成(包括整个复杂地球系统),应该采用哪种理论?这一系统的条件及空间尺度为何?目前,这些问题并没有完全解决。下文中,我们将对当前基于模型分析和真实系统中的空间斑图得出的理解进行概述,并明确指出系统中缺少的知识。
稀树草原生态系统的特征为树木和草地共存,大部分同质模型重现了这种共存关系。他们表明,根据降雨量、火灾强度和植被水平,开阔的稀树草原(以及分散的树木)通常可以在多种可选择的同质状态之间切换(见图4) ,其状态为:无树木;荒漠或草原;主要是稀树草原的封闭树木覆盖;稀树林地,或主要是森林树木的封闭树木覆盖;或热带雨林[5,10,11,38,64,65,75]。换句话说,忽略空间效应,这些模型预测,稀树草原生态系统可能呈现较次状态和临界转变以及两者间的临界。然而,当空间扩展时,这些模型通过图灵斑图[71,104]的出现和空间中可选稳定状态的共存而表现出丰富的空间斑图[51,73](见图4)。分析(Box1)指出,系统能否通过空间斑图规避临界点,都是有可能的。无论是热带雨林还是稀树草原,其恢复力通过与空间斑图形成相关的多稳态降低或是增加,都取决于局部但系统性的条件(如模型中的参数)。
临界行为和临界转变归因于许多地球系统的组成[3,15]。然而,即使对于这些系统来说,临界点的框架可能过于有限,而且多稳态的作用可能比以前认为的更重要。
作为一个经典的例子,我们考虑空间效应如何引入共存态,从而改变地球全球气候模型[52,53,105,106]中与雪冰反馈(ice-albedo feedback)相关的临界行为。地球温度的变化与能量收支的变化直接相关,能量收支的计算方法是入射太阳辐射减去反射和出射(普朗克)辐射。反射率的温度依赖性是雪冰反馈:只要温度低,地球就会被冰覆盖,冰面反射大部分太阳辐射。然而,当温度上升时,冰融化,冰面反射的辐射减少,从而导致温度进一步上升。如果不考虑空间效应,这种反馈机制在模型中会导致两种不同的地球状态:一个完全被冰覆盖的地球(”雪球地球”)或一个没有冰的地球(”无冰地球”)。存在一个双稳态区域,当某种稳态消失时,这两种状态之间的临界转变和临界点就出现了。
然而,在现实中,我们显然没有处于这两种地球状态中的任何一种,冰只存在于极地地区。这是因为该模型中,在全球能量预算中扮演重要作用的空间效应被忽略了。例如,入射太阳辐射与纬度有关,经向热流是能量分布的一个组成部分。当增加这种空间机制时,在雪球和无冰地球状态之外,模型还预测了冰和无冰的共存态。在共存态下,冰只存在于地球的一部分——靠近极点的地方有冰,同时赤道没有冰,这些可选状态之间的空间边界出现在某个中纬度。这些额外态的存在改变了非空间系统的经典临界属性:当一个完全(未)覆盖的地球状态消失时,向一个具有共存态系统的不那么临界的转变过程可能会发生,并且更多渐进的过渡是可能的。
除了这个共存态的例子之外,还有地球系统组件的多稳态。事实上,Busse balloon 起源于对热对流的研究,热对流[35]与流体力学中的湍流密切相关,因此也与全球大气和地球海洋环流密切相关。然而,在上面提到的大西洋环流经向翻转的例子中,当使用的盒子模型(box model,以连续性方程描述各组分速率不同的系统,常用于全球气候系统)仅具有相对较少的盒子时,可以观察到临界现象[107],而不能检测到共存态的多稳态。我们认为,尽管考虑了盒子之间的空间过程,若仅在有限数量的盒子内进行同质化,也会严重限制系统许多空间响应的涌现。通过增加盒子的数量来提高空间分辨率,增加了稳态的数量和性质[108,109],逐渐恢复了多稳态。在这方面,一个有趣的研究方向是调查地球系统组分的这种临界现象是否普遍[15]存在于高分辨率模型中(如最先进的全球气候模型[16] )或者被共存态的多稳定性(包括更多的渐进转变)所取代。如果是后者,这可能能够解释为什么复杂性更高的全球气候模型似乎比简单或中等复杂的模型[110]更稳定。
稀树草原的特点是草和树木共存,这种生态占据了超过八分之一的陆地面积,是很多动物的生存之源[60]。气候和土地使用的变化,会给全球稀树草原分布及植被特征带来显著变化[61,62]。树木覆盖的区域变动很大,且难以预测;有理论表明,空间斑图是导致该问题的主要因素,在很多对稀树草原[63]的研究中却是缺失的。稀树草原可以作为推导并验证能否规避临界点的生态系统模型,因为如下两个理由。
首先,来自模型和真实生态系统的观测指出,存在多个可选状态和临界点现象。生态系统可以在以下各状态(图4)间转变:无树木的生态系统(荒漠或草地),开阔稀树草原(草地上有零星树木),封闭稀树林地(覆盖着树木和灌木),和封闭热带雨林[5,8-12,38,64-68]。相比森林中的树木[69],稀树草原中的树木对火灾的恢复力较强,对处于荫蔽的恢复力较差。这些因素再结合草地易燃以及经历火灾后能轻易再次生长的特征,就是前文提到的正反馈的关键,形成了开阔稀树草原和热带雨林之间的多稳定性。第二,前文提到的两种空间斑图的生成,即图灵斑图和共存态,已在模型和真实生态系统中被观测到[12,25,68,70-76](图4)。因此,研究稀树草原生态系统为获取具有一般性价值和应用潜力的结果提供了一个绝佳机会。
当前研究表明,不存在一个统一的稀树草原模型能解释所有可能的生态系统模型(包括图灵斑图和共存态)。为了建立一个统一的、考虑空间斑图的稀树草原模型,可以通过如下方式整合现有的稀树草原模型[71,76,77],使新模型包含关于水、草地的生物量、稀树草原树木的生物量以及森林树木的生物量。草原灌木和森林树木的区别,在于面对火灾及处于荫蔽处的反应 [78, 79]。考虑到降雨、火灾、食草动物的非空间模型,会有助于创立这样一个统一的模型[64, 80],关于稀树草原-森林转变的研究也可能很有用 [10, 81]。新模型可以包含流入土壤和生物中的降水,以及火灾前后草地的生物量[11]。食草动物可以作为一个新的负反馈[82],或一个新的动力学方程加入[83]。之后该模型可被用来分析随着降雨梯度的变化而出现的下列问题:在什么条件下,开阔稀树草原和热带雨林之间,以及稀树草原和荒漠之间,会出现明显的分界线或渐进的空间边界?这些分界线如何根据气候、食草动物和火灾情况而演化、分岔及改变?在何种情况下,空间斑图会出现?出现的是图灵斑图还是共存态?可通过模拟、分岔分析及数值延拓来回答这些问题[84, 85]。
图4. 沿着降雨梯度,稀树草原的不同状态。依据年均降雨量[67]的高低,稀树草原通常可分为三类:湿润型、中湿型和干旱型(图中蓝带表示:从左到右,降雨量逐渐增大)。湿润区域可能是有热带雨林的双稳态;中湿区域可能是有稀树林地的双稳态;干旱区域则可能是有贫瘠荒漠的双稳态。在稀树草原状态的动态转变中,火与食草作用具有较大影响,它们共同决定了特定降雨条件下的树木覆盖度。从湿润区到干旱区,随着可用水的限制作用不断增强,火的重要性逐渐下降(图中红色条带)。研究将预期的空间结构与谷歌地球结果做了比较。自左向右,随着降雨量增大,图中分别展示了荒漠和干旱稀树草原的图灵斑图、稀树林地与中湿开阔稀树草原的共存态和图灵斑图,以及湿润稀树草原与森林的共存态。目前尚没有统一的模型可以解释所有观测状态与空间斑图。本文认为,未来有必要开发出一个统一的稀树草原模型。这一开发过程,需要纳入系统相关状态的变量与参数,同时结合数学分岔分析、模拟和数值延拓等多种方法[84, 85]。
基于这个统一模型的新的一般性预测可以通过如下方法验证(图4):
(i) 在降水梯度的湿润一端,在稀树草原-热带雨林的分界线,这里火灾情况通常被认为决定了开阔稀树草原和热带雨林这两个双稳态间切换的情况,我们预期会发现共存态以规避临界点[68, 86]。
(ii) 在降水梯度的中等湿润区,火灾和食草情况共同决定了封闭稀树林地和开阔稀树草原之间的双稳态切换,我们预期会出现共存态和图灵班图以规避临界点[70, 73, 74]。
(iii) 在降水梯度的干旱一端,通常是干旱决定了开阔稀树草原和荒漠的分界线,我们预期和Busse balloon 有关的图灵斑图导致了临界点的规避[33, 34]。
(iv) 火灾强度的增加会导致热带雨林-稀树草原的边界向热带雨林移动(湿润稀树草原取代森林)[81]。火灾强度或食草动物减少,会导致稀树林地取代中湿开阔稀树草原,而降雨的减少会导致荒漠取代干旱稀树草原。
在降雨梯度的湿润端,没有形成空间斑图的热带雨林-稀树草原的尖锐边界[87]可以解释为临界转变的空间替代[88]。然而,尚不清楚热带雨林-稀树草原边界在什么条件下是尖锐或渐进的,它们是否在移动,以及是否存在交替稳定状态(共存态)的斑图结构。因此,与上面描述的模型分析相同的问题可以通过观察来解决。经过模型分析的检验,上述预测可以得到验证,重点是火灾和干旱是主要的驱动因素。热带雨林和开阔稀树草原之间尖锐的植被边界[88]的空间特征可以通过卫星数据[87,89]在全世界范围内探索。这可以在多种空间分辨率下完成,创建多年或数十年[90]的时间序列,以便在筛选共存态[68,86]和图灵斑图的空间斑图结构边界所必需的较粗和较细的分辨率上探测植被边界,而这两者仅使用较粗的空间分辨率很容易被忽略。对于降雨梯度中间区的开阔稀树草原-稀树林地边界,对于干旱区的稀树草原-沙漠边界,也可以采用同样的方法。
通过变化指标[91,92],这些发现可能与主要驱动因素有关,如上面提到的干旱和火灾。空间边界的度量也可以与年度烧毁面积图[93]相关联。这将允许测试这些驱动因素如何决定植被边界的清晰度,共存态和图灵斑图的斑图结构是否出现,以及边界的移动如何与主要驱动因素的变化相关。考虑到气候和土地利用的预期变化,这一点很重要,而且它将使人们了解可能规避临界点的条件。
关于空间斑图形成的数学文献的主要部分集中在接近同质平衡的图灵斑图的出现[94]。然而,处于自然状态的真实生态系统和地球系统组分通常并不接近最初出现斑图的开端,而是远离平衡,空间斑图得到充分发展。对于实际系统中的许多空间斑图,只能有有限的数学洞察力,除非管理系统显示出足够大的尺度分离——这是系统各组成部分的空间传输在截然不同的空间和时间尺度上发挥作用的条件。在数学文献中,发生这种现象的系统被称为奇点扰动(singularly perturbed)。值得注意的是,这里所考虑的空间扩展模型和实际系统通常是奇点扰动的,奇点扰动的性质源于观测,即生态系统中的斑图形成实际上由广泛不同空间尺度上的反作用反馈机制驱动[31]。因此,这里所考虑的现实的、远离平衡的斑图可以用奇点扰动理论的方法在数学上详细研究[95]。此外,生态系统和地球系统组分通常显示的最相关斑图(包括Box2和3的例子)具有相互作用的局部结构的性质,例如植被带被裸露的土壤带包围的地区[33]、上文提到的开阔稀树草原和稀树林地的边界[73],以及冰川接地线[54]。多稳定性的恢复力[33,34]直接耦合于相互作用的局部结构(如局部条纹和空间边界)相关的丰富的各种模式。
因此,这些基本空间斑图和多稳定性的形成提供了一些机制,使空间生态系统(Box2)或地球系统组分(Box3)(包括整个复杂地球系统)一般可以规避临界。这种情况是否发生将取决于实际系统中的局部(但是系统性的)条件,或者取决于模型中的参数组合。对于不同类别的系统,将存在普遍的条件,导致通过空间斑图的形成和多稳定性规避临界点,例如稀树草原生态系统。这些是目前预测临界点的各类生态系统和地球系统组分。
对空间扩展的生态系统,如干旱稀树草原,用激活-抑制类型的反应-扩散方程[27,75,76]模拟的可靠性分析表明,临界之前可能有或可能没有形成斑图的图灵分岔(Box1和图1)。这种情况能否发生的条件可以用模型参数来明确表示(Box1)。然而,图灵分岔只能洞察空间斑图的开始,而不能洞察它们在开端之后(远离平衡)的行为,那时斑图持续发展并变得更容易观察。此外,正是这种开端之后的动力学决定了图灵斑图是否以及如何导致了对临界的规避。关于干旱稀树草原的文献[32,34]中已经阐明了一个明确的场景,其中图灵斑图的演化超越了起始阶段,在系统中规避了临界。现在的关键问题是,这种情况是否也可能在其他类型的系统中发挥作用,以及是否存在其他情况,通过这些情况,小幅度的图灵斑图可能演变为大规模的相互作用局部结构,使系统能够规避临界。由于模型的奇点扰动性质,可以用数学方法研究基本的局部结构及其相互作用[96-98]。为了揭示这些局部模式与图灵分岔出现的斑图之间的关键联系,有必要结合分岔分析、模拟和数值延拓[32,84,85]对 Busse balloon 进行进一步分析,特别是分析其边界的性质。
这样的分析也将揭示系统在分岔点前的动力学行为,从而导致更好的预警信号。经典的临界理论主要基于非空间模型,导致在临界发生之前产生一般的早期预警信号[2,99] ,主要与扰动后恢复的临界慢化(critical slowing)有关。在空间系统中,这个想法变得更加复杂,因为这现在可能关键取决于不同空间扰动的性质;临界慢化只显示具有特定空间结构的扰动[74] ,这可能很难甚至不可能在空间平均数据中检测到。同时,确实表现出临界慢化的扰动形式有助于确定系统发生前的分岔类型,即系统是否会通过形成空间斑图而达到或规避临界。当系统暴露于一般扰动(可能是随机扰动或局部扰动)之后,系统将在一个暂态恢复期内恢复,并可能暂时涌现斑图态或非斑图态。这种涌现的暂态(transient state)的形式现在可以是独特的,并且它所预示的分岔类型可以被识别。这种涌现的暂态导致图灵分岔,可能是一些空间周期斑图。相比之下,临界分岔之前可能会出现空间同质或极端局部的暂态。这样就有可能在空间时间序列发生之前就区分这两种类型的分岔。当然,一个主要的挑战是从数据中提取这些空间扰动,但是现有的所谓的模式分解算法[100]可能是一个可行的选择。
共存态之间的孤立界面,如开阔稀树草原-稀树林地边界、热带雨林-稀树草原边界(Box2和图4),以及海洋和冰之间的界面(Box3),对于多稳定性是必要的,通过多稳定性,整个空间系统由于稳定态扰动导致的临界得以避免。然而,在外部条件恶化的情况下,为了使空间共存斑图在超过临界点之后持续存在,可能需要一个额外的机制。正如同质状态被图灵分岔破坏一样,共存态之间的空间边界通常也可能分岔,从而可能成为多种演化中的局部空间斑图的起源。随后,这些模式可能会为生态系统或地球系统组分提供进一步的多稳定性,并使其可能沿着各种渐进路线超越临界点——也就是说,在环境条件恶化时规避临界点。这些分岔可以用数值方法来描述[51,55]。但更重要的是,模型的奇点扰动性质允许对潜在的不稳定机制进行基本理解[98,101]。例如,可以推导出这样的条件:离开均匀稳定状态的一个入侵边界会引发另一个斑图状态(其中包括多稳定性,即规避临界点的一条渐进路线)的反入侵。
在空间扩展的复杂系统中,大多数关于空间斑图形成的数学研究都假设一个高度理想化的区域——一个足够大的开放空间,环境条件在整个区域内不会改变。然而,在现实系统中,这种理想化的区域并不存在,而且结果是否以及如何从理想化的区域转移到更现实的区域也不明显。例如,如果一个系统可以演化的空间区域太小,无法形成空间斑图,那么规避临界就不再起作用。一个突出的例子可能是可选稳定态,和空间限制的浅水湖泊的临界[6]。这意味着,要使复杂系统形成空间斑图和增强恢复力需要一个最小区域面积;这一最小区域面积将取决于主要机制的空间尺度,和所考察的具体系统形成的空间斑图。
此外,局部效应(例如人为干预产生的效应)对斑图的形成、稳定性和动力学有很大影响[102]。这方面的例子包括砍伐热带雨林,固定沙丘或在沿海沙丘系统建造堤坝。这样的人为干扰尽管是局部的,也可能会大大降低灵活性,从而降低整个系统的恢复力。与之类似,也许更重要的是,如果人类将初始系统的空间斑图同化为非局部的,同样的情况也会发生,因为这里概述的增强恢复力的机制将不再起作用。这方面的例子包括陆地生态系统的大规模农业,防治荒漠化的空间同质性恢复努力,以及海洋生态系统的破坏性底拖网捕捞。因此,通过空间斑图形成来研究复杂系统的恢复力,应该深入分析区域空间限制的影响,以及局部和非局部同质化的人为诱发效应的影响。一个相关的方法将是结合计算和分析研究,以确定这种空间同质性对斑图动力学和恢复力的影响[103]。这对于恢复生态系统和减轻土地使用和气候变化的影响至关重要。
在这里,我们展示了图灵斑图产生的空间自组织和多稳定性如何帮助复杂系统规避临界点并增强恢复力。此外,我们概述了由于更全面的空间斑图形成和多稳定性,包括共存态的出现,规避临界点的条件和机制的范围应该比那些导致图灵斑图的范围要广得多。我们强调,这两种类型的空间斑图形成起源于临界点之前,并可以持续到临界点之后,展示了各种可以规避临界点的路径,同时强有力地增强了恢复力。我们强调,最近也观察到,这种空间斑图的形成和多稳定性适用于真正的生态系统,并认为,这可能与许多生态系统和地球系统组分有关(包括整个复杂的地球系统)。由于对临界现象和空间斑图形成的观察,热带草原可以作为一个典型生态系统来进一步研究这个问题。我们需要更好地理解空间斑图形成的一般动力学,以确定这些斑图如何响应不同程度和速率的外部变化,以及如何响应局部及非局部的同质扰动。这种认识将有助于确定哪些条件和空间斑图能够规避临界。我们预计,对许多生态系统和地球系统组分中临界现象的发现,将揭示一些系统实际上比目前认为的具有更大的恢复力。
Turing-before-tipping 临界转变前的图灵斑图
[1] M. Scheffer, S. Carpenter, J. A. Foley, C. Folke, B. Walker,Catastrophic shifts in ecosystems.Nature413, 591–596(2001). doi:10.1038/35098000; pmid:11595939
[2] M. Schefferet al., Early-warning signals for critical transitions.Nature461, 5 3–59 (2009). doi:10.1038/nature08227; pmid:19727193
[3] W. Steffenet al., Trajectories of the Earth System in the Anthropocene.Proc. Natl. Acad. Sci. U.S.A.115, 8252–8259 (2018). doi:10.1073/pnas.1810141115; pmid:30082409
[4] J. van de Koppel, M. Rietkerk, F. J. Weissing, Catastrophic vegetation shifts and soil degradation in terrestrial grazing systems.Trends Ecol. Evol.12, 352–356 (1997). doi: 10.1016/S0169-5347(97)01133-6; pmid:21238102
[5] M. Rietkerk, F. van den Bosch, J. van de Koppel, Site-specific properties and irreversible vegetation changes in semi-arid ecosystems.Oikos80, 241–252 (1997). doi:10.2307/ 3546592
[6] M. Scheffer, S. H. Hosper, M.-L. Meijer, B. Moss, E. Jeppesen,Alternative equilibria in shallow lakes.Trends Ecol. Evol.8,275–279 (1993). doi:10.1016/0169-5347(93)90254-M; pmid:21236168
[7] M. Rietkerk, J. van de Koppel, Alternate stable states and threshold effects in semi-arid grazing systems.Oikos79, 69–76 (1997). doi:10.2307/3546091
[8] M. Hirota, M. Holmgren, E. H. Van Nes, M. Scheffer, Global resilience of tropical forest and savanna to critical transitions.Science334, 232–235 (2011). doi:10.1126/ science.1210657; pmid:21998390
[9] A. C. Staver, S. Archibald, S. A. Levin, The global extent and determinants of savanna and forest as alternative biome states.Science334, 230–232 (2011). doi:10.1126/
science.1210465; pmid:21998389
[10] A. C. Staver, S. A. Levin, Integrating theoretical climate and fire effects on savanna and forest systems.Am. Nat.180, 211–224 (2012). doi:10.1086/666648; pmid:22766932
[11] F. Van Langeveldeet al., Effects of fire and herbivory on the stability of savanna ecosystems.Ecology84, 3 3 7–350 (2003). doi:10.1890/0012-9658(2003)084
[12] J. C. Alemanet al., Floristic evidence for alternative biome states in tropical Africa.Proc. Natl. Acad. Sci. U.S.A.117, 28183–28190 (2020). doi:10.1073/
pnas.2011515117;pmid:33109722
[13] C. S. Holling, inEngineering Within Ecological Constraints, P. E. Schulze, Ed. (National Academy Press, 1996), pp. 31–43.
[14] P. Ashwin, S. Wieczorek, R. Vitolo, P. Cox, Tipping points in open systems: Bifurcation, noise-induced and rate-dependent examples in the climate system.P h i l . T r a n s . R . S o c . A370, 1166–1184 (2012). doi:10.1098/rsta.2011.0306; p m i d :22291228
[15] T. M. Lentonet al., Tipping elements in the Earth’s climate system.Proc. Natl. Acad. Sci. U.S.A.105, 1786–1793 (2008). doi:10.1073/pnas.0705414105; pmid:18258748
[16] S. Drijfhoutet al., Catalogue of abrupt shifts in Intergovernmental Panel on Climate Change climate models. Proc. Natl. Acad. Sci. U.S.A.112, E5777–E5786 (2015). doi:10.1073/pnas.1511451112; pmid:26460042
[17] P. Huybrechts, J. de Wolde, The dynamic response of the Greenland and Antarctic ice sheets to multiple-century climatic warming.J. Clim.12, 2169–2188 (1999). doi:10.1175/
1520-0442(1999)012<2169:TDROTG>2.0.CO;2
[18] F. Pattyn, M. Morlighem, The uncertain future of the Antarctic ice sheet.Science367, 1331–1335 (2020). doi:10.1126/ science.aaz5487; pmid:32193321
[19] J. Garbe, T. Albrecht, A. Levermann, J. F. Donges, R. Winkelmann, The hysteresis of the Antarctic ice sheet. Nature585, 538–544 (2020). doi:10.1038/s41586-020- 2727-5; pmid:32968257
[20] T. F. Stocker, D. G. Wright, Rapid transitions of the ocean’s deep circulation induced by changes in surface water fluxes. Nature351, 729–732 (1991). doi:10.1038/351729a0
[21] S. Rahmstorf, A. Ganopolsky, Long-term global warning scenario’s computed with an efficient coupled climate model. Clim. Change43, 353–367 (1999). doi:10.1023/
[22] J. Lohmann, P. D. Ditlevsen, Risk of tipping the overturning circulation due to increasing rates of ice melt.Proc. Natl. Acad. Sci. U.S.A.118, e20179891 (2021). doi:10.1073/pnas.2017989118; pmid:33619095
[23] J. Rockströmet al., A safe operating space for humanity. Nature461, 472–475 (2009). doi:10.1038/461472a; pmid:19779433
[24] A. D. Barnoskyet al., Approaching a state shift in Earth’s biosphere.Nature486, 5 2–58 (2012). doi:10.1038/ nature11018; pmid:22678279
[25] M. Rietkerk, S. C. Dekker, P. C. de Ruiter, J. van de Koppel, Self-organized patchiness and catastrophic shifts in ecosystems.Science305, 1926–1929 (2004). doi:10.1126/ science.1101867; pmid:15448261
[26] A. M. Turing, The chemical basis of morphogenesis. Bull. Math. Biol.52, 153–197 (1990). doi:10.1007/ BF02459572; pmid:2185858
[27] C. A. Klausmeier, Regular and irregular patterns in semiarid vegetation.Science284, 1826–1828 (1999). doi:10.1126/ science.284.5421.1826; pmid:10364553
[28] J. von Hardenberg, E. Meron, M. Shachak, Y. Zarmi, Diversity of vegetation patterns and desertification.Phys. Rev. Lett.87, 198101 (2001). doi:10.1103/
[29] M. Rietkerket al., Self-organization of vegetation in arid ecosystems.Am. Nat.160, 524–530 (2002). doi:10.1086/ 342078; pmid:18707527
[30] K. Gowda, H. Riecke, M. Silber, Transitions between patterned states in vegetation models for semiarid ecosystems. Phys. Rev. E89, 022701 (2014). doi:10.1103/ PhysRevE.89.022701; pmid:25353503
[31] M. Rietkerk, J. van de Koppel, Regular pattern formation in real ecosystems.Trends Ecol. Evol.23, 169–175 (2008). doi:10.1016/j.tree.2007.10.013; pmid:18255188
[32] K. Siteuret al., Beyond Turing: The response of patterned ecosystems to environmental change.Ecol. Complex.20, 81–96 (2014). doi:10.1016/
[33] R. Bastiaansenet al., Multistability of model and real dryland ecosystems through spatial self-organization.Proc. Natl. Acad. Sci. U.S.A.115, 11256–11261 (2018). doi:10.1073/ pnas.1804771115; pmid:30322906
[34] R. Bastiaansen, A. Doelman, M. B. Eppinga, M. Rietkerk, The effect of climate change on the resilience of ecosystems with adaptive spatial pattern formation.Ecol. Lett.23, 4 1 4–429 (2020). doi:10.1111/ele.13449; pmid:31912954
[35] F. H. Busse, Non-linear properties of thermal convection. Rep. Prog. Phys.41, 1929–1967 (1978). doi:10.1088/ 0034-4885/41/12/003
[36] R. H. Wang, Q. X. Liu, G. Q. Sun, Z. Jin, J. van de Koppel, Nonlinear dynamic and pattern bifurcations in a model for spatial patterns in young mussel beds.J. R. Soc. Interface6, 705–718 (2009). doi:10.1098/rsif.2008.0439; pmid:18986965
[37] Q. X. Liuet al., Pattern formation at multiple spatial scales drives the resilience of mussel bed ecosystems.Nat. Commun. 5, 5234 (2014). doi:10.1038/ncomms6234; pmid:25335554
[38] E. H. van Nes, M. Hirota, M. Holmgren, M. Scheffer, Tipping points in tropical tree cover: Linking theory to data. Glob. Change Biol.20, 1016–1021 (2014). doi:10.1111/ gcb.12398; pmid:24106057
[39] J. Pastor, B. Peckham, S. Bridgham, J. Weltzin, J. Chen, Plant community dynamics, nutrient cycling, and alternative stable equilibria in peatlands.Am. Nat.160, 553–568 (2002). doi:10.1086/342814; pmid:18707507
[40] M. J. Donahue, R. A. Desharnais, C. D. Robles, P. Arriola, Mussel bed boundaries as dynamic equilibria: Thresholds, phase shifts, and alternative states.Am. Nat.178, 612–625 (2011). doi:10.1086/662177; pmid:22030731
[41] J. Van de Koppel, P. M. J. Herman, P. Thoolen, C. H. R. Heip, Do alternate stable states occur in natural ecosystems? Evidence from a tidal flat.Ecology82, 3449–3461 (2001). doi:10.1890/0012-9658(2001)082[3449:DASSOI]2.0.CO;2
[42] O. D. Vinent, L. J. Moore, Barrier island bistability induced by biophysical interactions.Nat. Clim. Chang.5, 158–162 (2015). doi:10.1038/NCLIMATE2474
[43]T. Van der Heide et al., Positive feedbacks in seagrass ecosystems: Implications for success in conservation and restoration. Ecosystems 10, 1311–1322 (2007). doi: 10.1007/ s10021-007-9099-7
[44] T. van der Heide et al., Spatial self-organized patterning in seagrasses along a depth gradient of an intertidal ecosystem. Ecology 91, 362–369 (2010) doi: 10.1890/
08-1567.1; pmid: 20392001
[45] E. D. Lazarus, S. Armstrong, Self-organized pattern formation in coastal barrier washover deposits. Geology 43, 363–366 (2015). doi: 10.1130/G36329.1
[46] D. Ruiz-Reynés et al., Fairy circle landscapes under the sea.
Sci. Adv. 3, e1603262 (2017). doi: 10.1126/sciadv.1603262;
[47] V. Reijers, “A song of sand and mud: How plant-mediated feedbacks dictate landscape formation and dynamics of barrier islands,”thesis, Radboud University, Nijmegen (2019).
[48] P. D. L. Ritchie, J. J. Clarke, P. M. Cox, C. Huntingford, Overshooting tipping point thresholds in a changing climate.Nature 592, 517–523 (2021). doi: 10.1038/s41586-021-
[49] H. Hillebrand et al., Thresholds for ecological responses to global change do not emerge from empirical data. Nat. Ecol. Evol. 4, 1502–1509 (2020). doi: 10.1038/s41559-020-1256-9;pmid: 32807945
[50] G. Bel, A. Hagberg, E. Meron, Gradual regime shifts in spatially extended ecosystem. Theor. Ecol. 5, 591–604 (2012). doi: 10.1007/s12080-011-0149-6
[51] Y. R. Zelnik, E. Meron, Regime shifts by front dynamics.Ecol. Indic. 94, 544 –552 (2018). doi: 10.1016/j.ecolind.2017.10.068
[52] M. Ghil, Climate stability for a Sellers-type model. J. Atmos. Sci. 33, 3 –20 (1976). doi: 10.1175/1520-0469(1976)033<0003:CSFAST>2.0.CO;2
[53] T. Bódai, V. Lucarini, F. Lunkeit, R. Boschi, Global instability in the Ghil–Sellers model. Clim. Dyn. 44, 3361–3381 (2015).doi: 10.1007/s00382-014-2206-5
[54] S. H. R. Rosier et al., The tipping points and early-warning indicators for Pine Island Glacier, West Antarctica. Cryosphere 15, 1501–1516 (2021). doi: 10.5194/tc-15-1501-2021
[55] C. Fernandez-Oto, O. Tzuk, E. Meron, Front instabilities can reverse desertification. Phys. Rev. Lett. 122, 048101 (2019).doi: 10.1103/PhysRevLett.122.048101; pmid: 30768298
[56] M. E. Cates, J. Tailleur, Motility-induced phase separation. Annu. Rev. Condens. Matter Phys. 6, 219–244 (2015).doi: 10.1146/annurev-conmatphys-031214-014710
[57] Q.-X. Liu et al., Phase separation explains a new class of self-organized spatial patterns in ecological systems. Proc. Natl. Acad. Sci. U.S.A. 110, 11905–11910 (2013). doi: 10.1073/pnas.1222339110; pmid: 23818579
[58] Q.-X. Liu et al., Phase separation driven by density-dependent movement: A novel mechanism for ecological patterns. Phys. Life Rev. 19, 107–121 (2016). doi: 10.1016/
j.plrev.2016.07.009; pmid: 27478087
[59] E. Sheffer, J. von Hardenberg, H. Yizhaq, M. Shachak, E. Meron, Emerged or imposed: A theory on the role of physical templates and self-organisation for vegetation patchiness. Ecol. Lett. 16, 127–139 (2013). doi: 10.1111/ele.12027; pmid: 23157578
[60] R. J. Scholes, S. R. Archer, Tree-grass interactions in savannas. Annu. Rev. Ecol. Syst. 28, 517–544 (1997).doi: 10.1146/annurev.ecolsys.28.1.517
[61] J. D. Anadon, O. E. Sala, F. T. Maestre, Climate change will increase savannas at the expense of forests and treeless vegetation in tropical and subtropical Americas. J. Ecol. 102, 1363–1373 (2014). doi: 10.1111/1365-2745.12325
[62] M. Garcia Criado, I. H. Meyers-Smith, A. D. Bjorkman, C. E. R. Lehman, N. Stevens, Woody plant encroachment intensifies under climate change across tundra and savanna biomes. Glob. Ecol. Biogeogr. 29, 925–943 (2020). doi: 10.1111/geb.13072
[63] A. C. Staver, Prediction and scale in savanna ecosystems. New Phytol. 219, 52 –57 (2018). doi: 10.1111/nph.14829;pmid: 29027662
[64] F. Accatino, C. De Michele, R. Vezzoli, D. Donzelli, R. J. Scholes, Tree–grass co-existence in savanna: Interactions of rain and fire. J. Theor. Biol. 267, 235 –242 (2010). doi: 10.1016/ j.jtbi.2010.08.012; pmid: 20708629
[65] M. Baudena, F. D’Andrea, A. Provenzale, An idealized model for tree-grass coexistence in savannas: The role of life stage structure and fire disturbances. J. Ecol. 98, 74 –80 (2010). doi: 10.1111/j.1365-2745.2009.01588.x
[66] V. de L. Dantas, M. Hirota, R. S. Oliveira, J. G. Pausas, Disturbance maintains alternative biome states. Ecol. Lett. 19, 12–19 (2016). doi: 10.1111/ele.12537; pmid: 26493189
[67] D. D’Onofrio, J. von Hardenberg, M. Baudena, Not only trees: Grasses determine African tropical biome distributions via water limitation and fire. Glob. Ecol. Biogeogr. 27, 714 –725 (2018). doi: 10.1111/geb.12735
[68] A. W. Cardoso et al., A distinct ecotonal tree community exists at central African forest-savanna transitions. J. Ecol.109, 1170–1183 (2021). doi: 10.1111/1365-2745.13549
[69] T. Charles-Dominique, G. F. Midgley, K. W. Tomlinson, W. J. Bond, Steal the light: Shade vs fire adapted vegetation in forest-savanna mosaics. New Phytol. 218, 1419–1429 (2018). doi: 10.1111/nph.15117; pmid: 29604213
[70] T. A. Groen,“Spatial matters: How spatial patterns and processes affect savanna dynamics,”thesis, Wageningen University (2007).
[71] M. Baudena, M. Rietkerk, Complexity and coexistence in a simple spatial model for arid savanna ecosystems. Theor. Ecol.6, 131 –141(2013).doi:10.1007/s12080-012-0165-1
[72] R. Martínez-García, J. M. Calabrese, C. López, Spatial patterns in mesic savannas: The local facilitation limit and the role of demographic stochasticity. J. Theor. Biol. 333,
156–165 (2013). doi: 10.1016/j.jtbi.2013.05.024; pmid: 23747988
[73] T. A. Groen, C. A. D. M. van de Vijver, F. van Langevelde, Do spatially homogenizing and heterogenizing processes affect transitions between alternative stable states? Ecol. Modell. 65, 119 –128 (2017). doi: 10.1016/j.ecolmodel.2017.10.002
[74] S. Eby, A. Agrawal, S. Majumder, A. P. Dobson, V. Guttal, Alternative stable states and spatial indicators of critical slowing down along a spatial gradient in a savanna
ecosystem. Glob. Ecol. Biogeogr. 26, 638–649 (2017).doi: 10.1111/geb.12570
[75] L. Eigentler, J. A. Sherratt, Metastability as a coexistence mechanism in a model for dryland vegetation patterns. Bull. Math. Biol. 81, 2290–2322 (2019). doi: 10.1007/ s11538-019-00606-z; pmid: 31012031
[76] O. Tzuk, H. Uecker, E. Meron, The role of spatial self-organization in the design of agroforestry systems.PLOS ONE 15, e0236325 (2020). doi: 10.1371/ journal.pone.0236325; pmid: 32692773
[77] B. Wuyts, A. R. Champneys, N. Verschueren, J. I. House, Tropical tree cover in a heterogeneous environment:A reaction-diffusion model. PLOS ONE 14, e0218151 (2019).doi: 10.1371/journal.pone.0218151; pmid: 31246968
[78] V. Yatat, P. Couteron, J. J. Tewa, S. Bowong, Y. Dumont, An impulsive modelling framework of fire occurrence in a size-structured model of tree-grass interactions for savanna ecosystems. J. Math. Biol. 74, 1425–1482 (2017). doi: 10.1007/s00285-016-1060-y; pmid: 27659304
[79] V. Yatat, P. Couteron, Y. Dumont, Spatially explicit modelling of tree–grass interactions in fire-prone savannas: A partial differential equations framework. Ecol. Complex. 36, 290–313(2018). doi: 10.1016/j.ecocom.2017.06.004
[80] C. De Michele, F. Accatino, R. Vezzoli, R. J. Scholes, Savanna domain in the herbivores-fire parameter space exploiting a tree–grass–soil water dynamic model. J. Theor. Biol. 289, 74 –82 (2011). doi: 10.1016/j.jtbi.2011.08.014;pmid: 21875600
[81] Q. Li, A. C. Staver, E. Weinan, S. A. Levin, Spatial feedbacks and the dynamics of savanna and forest. Theor. Ecol. 12, 237–262 (2019). doi: 10.1007/s12080-019-0428-1
[82] E. Siero et al., Grazing away the resilience of patterned ecosystems. Am. Nat. 193, 472–480 (2019). doi: 10.1086/701669; pmid: 30794443
[83] R. M. Holdo, R. D. Holt, J. M. Fryxell, Herbivore-vegetation feedbacks can expand the range of savanna persistence:Insights from a simple theoretical model. Oikos 122, 441–453(2013). doi: 10.1111/j.1600-0706.2012.20735.x
[84] J. Sherratt, Numerical continuation methods for studying periodic travelling wave (wavetrain) solutions of partial differential equations. Appl. Math. Comput. 218, 4684–4694(2012). doi: 10.1016/j.amc.2011.11.005
[85] S. Van der Stelt, A. Doelman, G. M. Hek, J. Rademacher, Rise and fall of periodic patterns for a generalized Klausmeier–Gray–Scott model. J. Nonlinear Sci. 23, 39 –95 (2013).doi: 10.1007/s00332-012-9139-0
[86] A. Staal, S. C. Dekker, C. Xu, E. Van Nes, Bistability, spatial interactions and the distribution of tropical forests and savannas. Ecosystems 19, 1080–1091 (2016). doi: 10.1007/s10021-016-0011-1
[87] J. C. Aleman, A. C. Staver, Spatial patterns in the global distributions of savanna and forest. Glob. Ecol. Biogeogr. 27, 792–803 (2018). doi: 10.1111/geb.12739
[88] D. R. Uden et al., Spatial imaging and screening for regime shifts. Front. Ecol. Evol. 7, 407 (2019). doi: 10.3389/fevo.2019.00407
[89] A. C. Staver, G. P. Asner, I. Rodriguez-Iturbe, S. A. Levin, I. P. J. Smit, Spatial patterning among savanna trees in high-resolution, spatially extensive data. Proc. Natl. Acad. Sci. U.S.A. 116, 10681–10685 (2019). doi: 10.1073/ pnas.1819391116; pmid: 31085650
[90] W. Zhang et al., From woody cover to woody canopies: How Sentinel-1 and Sentinel-2 data advance the mapping of woody plants in savannas. Remote Sens. Environ. 234, 111465(2019). doi: 10.1016/j.rse.2019.111465
[91] G. Xanthopoulos, G. Maheras, V. Gouma, M. Gouvas, Is the Keetch–Byram drought index (KBDI) directly related to plant water stress? For. Ecol. Manage. 234, S27 (2006).
doi: 10.1016/j.foreco.2006.08.043
[92] L. Wang, J. J. Qu, Satellite remote sensing applications for surface soil moisture monitoring: A review. Front. Earth Sci. China 3, 237 –247 (2009). doi: 10.1007/s11707-009-0023-7
[93] T. Long et al., 30 m Resolution Global Annual Burned Area Mapping Based on Landsat Images and Google Earth Engine. Remote Sens. 11, 489 (2019). doi: 10.3390/rs11050489
[94] R. B. Hoyle, Pattern Formation: An Introduction to Methods (Cambridge Univ. Press, 2006).
[95] A. Doelman, in Complexity Science, M. Peletier, R. van Santen, E. Steur, Eds. (World Scientific, 2019), pp. 129–192.
[96] R. Bastiaansen, A. Doelman, The dynamics of disappearing pulses in a singularly perturbed reaction-diffusion systems with parameters that vary in time and space. Physica D 388, 45–72 (2019). doi: 10.1016/j.physd.2018.09.003
[97] R. Bastiaansen, P. Carter, A. Doelman, Stable planar vegetation stripe patterns on sloped terrain in dryland ecosystems. Nonlinearity 32, 2759–2814 (2019).
doi: 10.1088/1361-6544/ab1767
[98] O. Jaïbi, A. Doelman, M. Chirilus-Bruckner, E. Meron, The existence of localized vegetation patterns in a systematically reduced model for dryland vegetation. Physica D 412, 132637 (2020). doi: 10.1016/j.physd.2020.132637
[99] M. Scheffer et al., Anticipating critical transitions. Science 338, 344–348 (2012). doi: 10.1126/science.1225244; pmid: 23087241
[100] J. N. Kutz, S. L. Brunton, B. W. Brunton, J. L. Proctor,Dynamic Mode Decomposition: Data-Driven Modeling of Complex Systems (Society for Industrial and Applied Mathematics, 2016).
[101] P. Van Heijster, B. Sandstede, Bifurcations to travelling planar spots in a three-component FitzHugh–Nagumo system. Physica D 275, 19 –34 (2014). doi: 10.1016/
[102] B. Wuyts, A. R. Champneys, J. I. House, Correction: Author Correction: Amazonian forest-savanna bistability and human impact. Nat. Commun. 9, 16179 (2018). doi: 10.1038/ncomms16179; pmid: 29465082
[103] A. Doelman, P. van Heijster, J. Shen, Pulse dynamics in reaction–diffusion equations with strong spatially localizedimpurities. Phil. Trans. R. Soc. A 376, 20170183 (2018).doi: 10.1098/rsta.2017.0183; pmid: 29507168
[104] L. Eigentler, J. A. Sherratt, Spatial self-organisation enables species coexistence in a model for savanna ecosystems.J. Theor. Biol. 487, 110122 (2020). doi: 10.1016/
j.jtbi.2019.110122; pmid: 31862451
[105] M. I. Budyko, The effect of solar radiation variations on the climate of the Earth. Tellus 21, 611–619 (1969). doi: 10.3402/tellusa.v21i5.10109
[106] W. D. Sellers, A global climatic model based on the energy balance of the earth-atmosphere system. J. Appl. Meteorol. 8,392–400 (1969). doi: 10.1175/1520-0450(1969)008<0392:AGCMBO>2.0.CO;2
[107] H. Stommel, Thermohaline convection with two stable regimes of flow. Tellus 13, 224–230 (1961). doi: 10.1111/j.2153-3490.1961.tb00079.x
[108] H. Gildor, E. Tziperman, Physical mechanisms behind biogeochemical glacial-interglacial CO2 variations.Geophys. Res. Lett. 28, 2421 –2424 (2001). doi: 10.1029/
[109] H. M. Alkhayoun, P. Ashwin, L. C. Jackson, C. Quinn, R. A. Wood, Basin bifurcation, oscillatory instability and scale-induced thresholds for Atlantic meridonal overturning
circulation in a global box model. Proc. R. Soc. A 475,20190051 (2019). doi: 10.1098/rspa.2019.0051
[110]. P. Valdes, Built for stability. Nat. Geosci. 4, 414 –416 (2011).doi: 10.1038/ngeo1200
集智斑图顶刊论文速递栏目上线以来,持续收录来自Nature、Science等顶刊的最新论文,追踪复杂系统、网络科学、计算社会科学等领域的前沿进展。现在正式推出订阅功能,每周通过微信服务号「集智斑图」推送论文信息。扫描下方二维码即可一键订阅:

点击“阅读原文”,追踪复杂科学顶刊论文