组织生物学:寻找新范式
摘要

论文题目:Tissue Biology: In Search of a New Paradigm
论文链接:
https://www.annualreviews.org/content/journals/10.1146/annurev-cellbio-120420-113830
组织通常被定义为具有共同形态和功能的细胞的集合。传统观点认为,动物组织主要有4种类型:上皮组织、结缔组织、肌肉组织和神经组织。然而,在许多情况下,组织更普遍地被认为是不同细胞类型的有组织的集合体。随着近年来组织生物学研究的增多,该领域的一些基本概念需要从现代角度进行重新架构,使其具有内在一致性、可推广性和知识性。这将有助于解决组织结构的一些基本问题:组织结构的设计原则是什么?不同的组织是否代表在一个共同设计主题上的变异(类似于不同的细胞类型是在真核细胞的基本设计上的变异)?组织中是否存在某种细胞类型的层次结构?如果是,其依据是什么?
一个与组织生物学特别相关的视角来自于复杂系统理论。复杂系统被定义为按照某种规则相互作用的多样的、相互依赖的、相互关联的主体的集合。这些相互作用的结果是系统涌现的性质 (其结构,功能,或动态行为),这些都是个体的特征所不能还原的。组织具有复杂系统的所有特征:它们是由不同的、相互联系的、相互依赖的细胞类型组成的,这些细胞类型按照一定的规则相互作用,从而产生组织结构、功能和组成的新特性。
1. 动物组织的进化过程
1. 动物组织的进化过程
探索更现代的组织和身体构成需要对先前存在的组织类型的框架进行阐述。追溯动物组织类型的演化,需要推断后生动物祖先(所有后生动物的假定最近共同祖先)的组织构成以及后生动物演化的早期事件。
后生动物包括5个主要谱系:两侧对称动物(Bilateria)、刺胞动物(Cnidaria)、栉水母类(Ctenophora)、扁盘动物(Placozoa)、海绵动物(Porifera)。两侧对称动物和刺胞动物为姐妹谱系,但其余群系间的系统发育关系尚未完全解析清楚。上皮层在5个谱系的动物中都存在 (图1),表明它是一种基本的后生动物组织,很可能存在于后生动物祖先中。除了基本的上皮组织外,所有动物中都存在一种间充质细胞类型:例如海绵中的原始细胞 (archaeocytes),扁盘动物门中的纤维细胞 (fiber cells)和脊椎动物中的成纤维细胞(fibroblasts)。最近的研究表明,领鞭毛虫在应激状态下转变为似变形虫样的状态。这表明在领鞭毛虫存在间充质表型,以及上皮-间充质转化 (EMT, epithelial-mesenchymal transition) 的机制在原后生动物(Urmetazoan)已经存在。上皮和间充质细胞的古老起源进一步支持了上皮-间充质单位在动物组织中的中心地位。
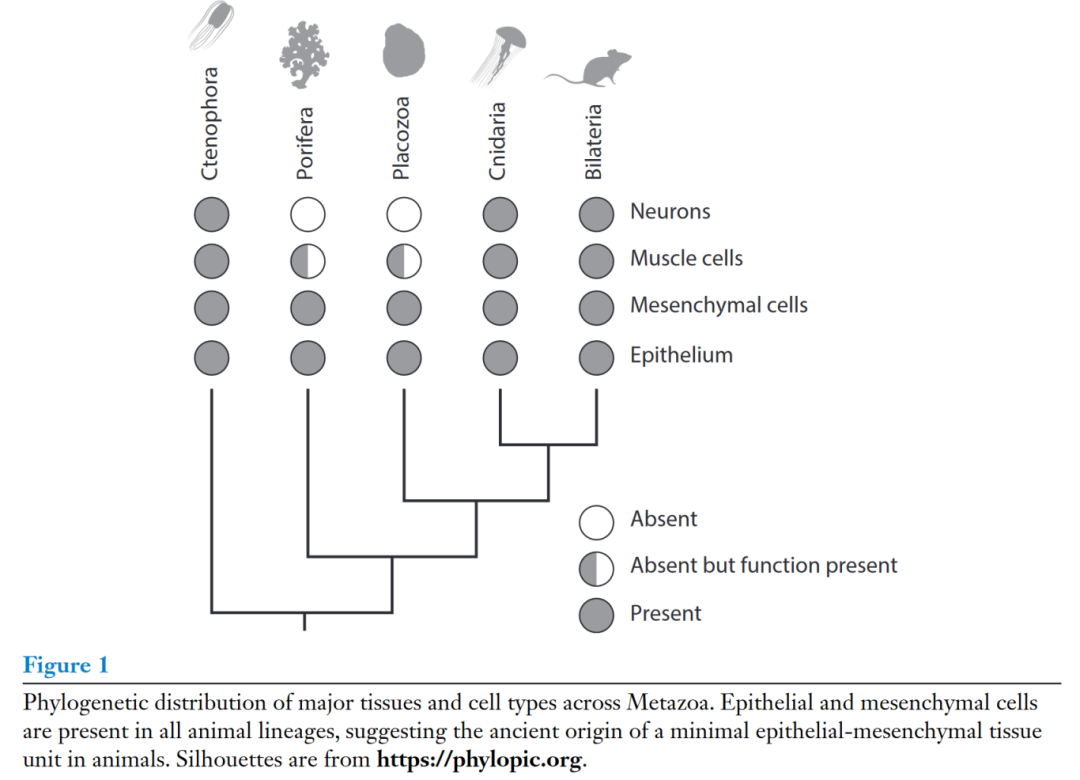
在上皮-间充质单元之外,栉水母类、刺胞动物和两侧对称动物具有特化的肌细胞,而海绵动物和扁盘动物门缺乏特化的肌细胞,但其有部分细胞类型为收缩型。因此,虽然肌细胞不是动物共有的特征,但收缩装置却是。在海绵中已经鉴定出表达神经元模块的细胞类型,如突触前和突触后机械;并且在扁盘动物门和中鉴定出可能调节摄食和运动的肽能神经分泌细胞。这些观察结果共同暗示了动虽然后生动物祖先缺乏神经系统,但它具有功能和调节模块,为栉水母类、刺胞虫和两侧对称动物的神经系统创造了先决条件。
总而言之,后生动物祖先的身体平面很可能被上皮层所束缚,或许在上皮层之间的空间中具有外形不规则的间充质细胞类型。上皮组织可能是多功能的,具有收缩、感觉和调节功能。
2. 细胞类型及其演化
2. 细胞类型及其演化
推测的原初动物祖先的组织组成表明,基本的组织类型和细胞类型在动物进化早期出现,随后经历了谱系特异性的扩张。组织类型的逐渐扩大很可能是通过细胞类型的多样化以及它们之间相互作用的演变而发生的。后生动物谱系中形态复杂性随时间的增加并不一定反映功能复杂性的增加。对两者进行明确的区分可以概念化一个细胞类型多样化的模型,其过程分为两步:在现有细胞类型(功能复杂性的增加)的基础上增加一个新的功能,然后将功能分离为两个姐妹细胞类型(形态复杂性的增加)。
下面这个模型概括了一个细胞类型可以演化为两个姐妹细胞类型的两种模式。这两种模式可以概括为从诱导型到组成型的转变(图2a)和分工的权衡(图2b)。在第一种模式中,新功能是由环境诱导的基因表达程序驱动的,该基因表达程序既可以是化学信号,也可以是基于细胞解剖位置的位置信号。起初,这种诱导状态可以是可逆的激活或极化状态。随后,诱导的基因表达程序可以演化为组成型。这类似于遗传同化作用(genetic assimilation),可以通过将诱导转录因子的表达置于谱系决定转录因子的控制之下来实现。
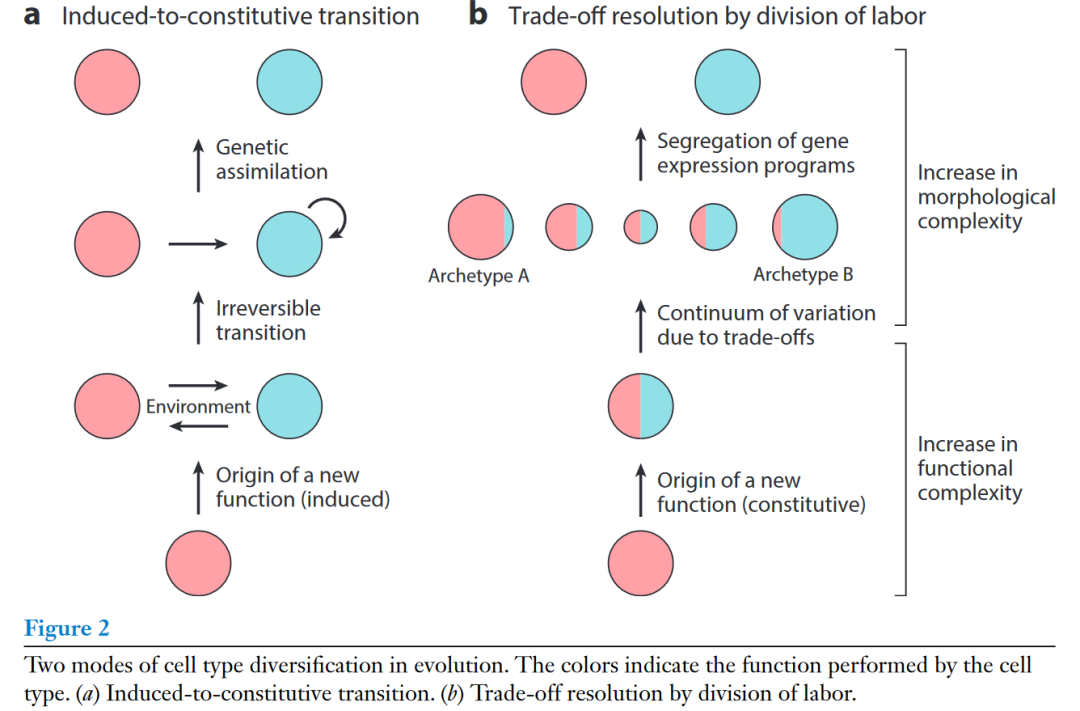
在第二种模式中,新功能被添加到祖先细胞类型中,作为组成型表达的新功能模块。因此,祖先细胞类型的两种功能并不是时间或空间上划定的,而是由同一细胞执行的。根据功能的性质,这种多功能性可以导致两种功能之间的权衡,使得单个细胞可以有效地执行一种功能,而只牺牲另一种功能。从而导致给定类型的细胞之间存在一个连续的变异 (在功能和基因表达方面)。这种细胞类型的个体在基因表达空间中占据的连续空间的顶点或极端位置代表了给定细胞类型的所谓原型(archetypes),这个原型相较于另一个功能更优先于现在的这个功能。随着时间的推移,可以通过夸大变异的连续体、关闭一个功能的基因表达程序和通过选择性保留另一个功能的基因表达程序来解决权衡。
3. 基于细胞间关系的细胞分类
3. 基于细胞间关系的细胞分类
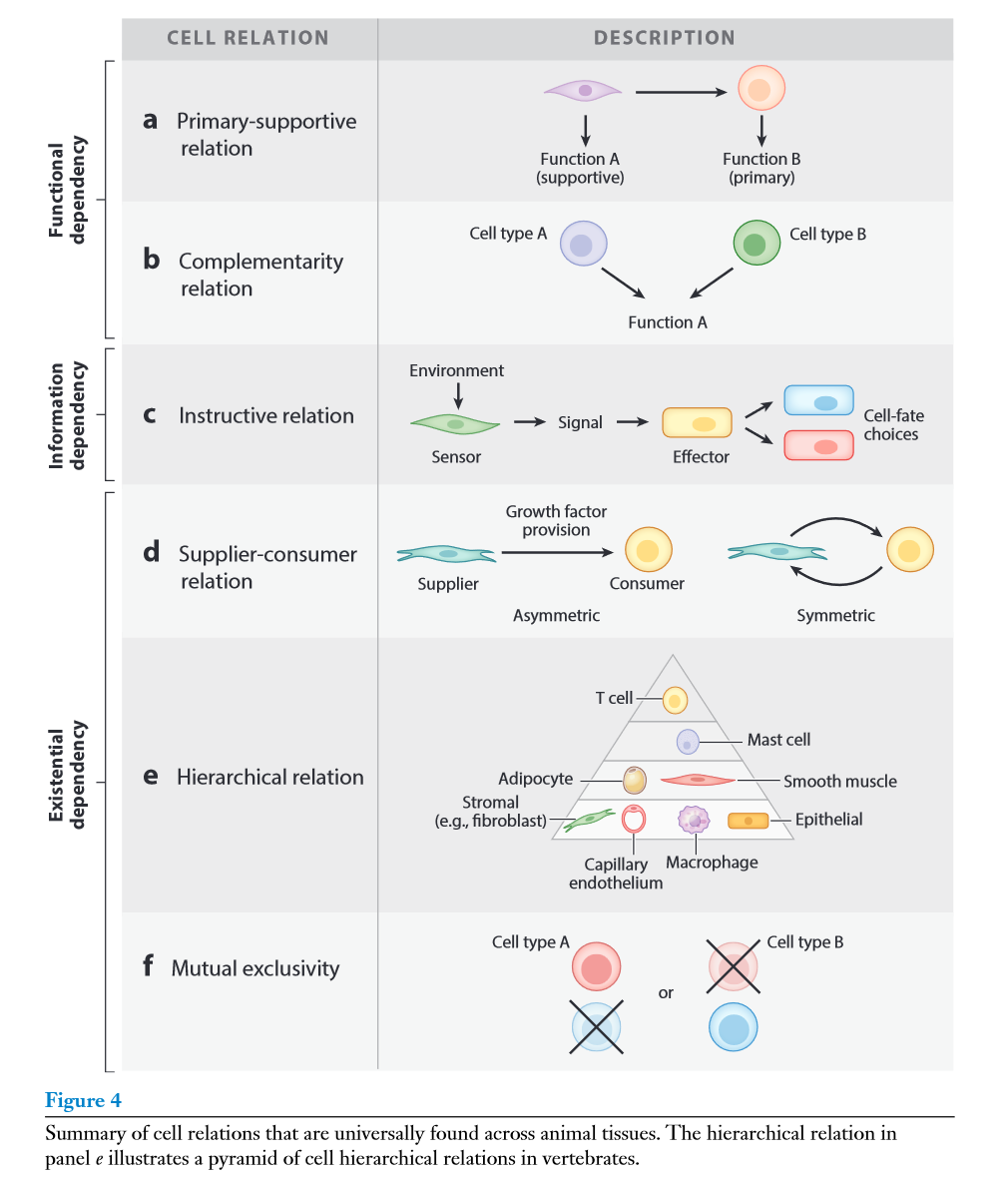
细胞类型的概念反映了细胞的内在特征,包括细胞的发育起源、功能和形态。为了在他们的社会背景中理解细胞,我们需要一个互补的特征来反映细胞之间的关系模式。关系定义了不一定与其身份(或细胞类型)相对应的细胞类别。基于身份和关系的分类的差异可以用社会系统作为类比来说明:仅仅使用关于个体特征的信息(例如,姓名、年龄、性别和职业) 来理解社会群体的行为是有限的。了解不同个体之间的(例如,亲子关系、雇主-雇员关系、配偶关系、朋友关系等)关系对理解社会结构至关重要。这些关系定义了配偶、父母、雇员或朋友的类别,它们提供了对一个社会群体的组织的洞察力,而这是仅仅从个体特征的知识中无法获得的。同样,基于单个细胞类型特征来表征细胞对于理解其在组织中的作用是不够的。为了在他们的社会背景中探索细胞,我们必须定义细胞类别,其中定义类别的不是细胞身份,而是它与其他细胞的关系(图3)。下面讨论了几种常见的细胞关系及其在组织中的作用:

3.1 执行-支撑关系
考虑到组织的功能组织性,细胞类型的多样性通常可以分为两类功能:执行组织主要功能的细胞和执行支持功能的细胞,其中支持细胞用于促进和优化主要组织功能的性能 (图4a)。原始-支持细胞关系在动物进化早期就出现在原始上皮-间充质组织单元中,其中上皮细胞执行防御、营养获取和维持内部稳态的主要功能,而间充质细胞通过产生细胞外基质(extracellular matrix,ECM) 和其他支持上皮细胞主要功能的分泌因子提供结构和功能支持。组织特异性功能由在不同组织中存在差异的原始细胞类型来执行,而支持细胞既可以是特殊的 (例如大脑中的胶质细胞或血管中的周细胞) ,也可以是大多数组织的通用细胞。后者包括成纤维细胞、毛细血管内皮细胞和组织驻留巨噬细胞。
3.2 互补关系
上面所描述的“执行-支撑”关系是非对称关系的一个例子:A细胞对B细胞是支持的,但反之则不然。然而,存在组织中的功能分工导致细胞同样贡献一个主要功能的情况——它们通过形成功能单元来互补彼此的功能 (图4b)。例如,成骨细胞和破骨细胞在基质沉积和重吸收中具有互补功能,成纤维细胞和巨噬细胞在总体来说也具有互补功能。
3.3 指导关系
原始上皮-间充质组织单元定义了另一种依赖于不对称信息传递的细胞关系。在胚胎诱导过程中,间充质细胞产生指令,如骨形态发生蛋白 ( bone morphogenetic protein,BMP) 抑制剂和成纤维细胞生长因子 (fibroblast growth factors,FGFs) 作用于Wnt响应的表皮细胞,使其分化为基板(placodes)。在这里,真皮细胞包含位置信息,而表皮细胞则有多个命运选择 (例如,形成不同的皮肤附属物)。表皮细胞的命运选择是由真皮细胞发出的信号决定的,而真皮细胞的信号又是由位置信息 (特异性Hox基因在真皮细胞中的表达) 决定的。这个例子说明了一个指导性的关系:细胞A具有决定细胞B命运选择的信息 (图4c),无论是A细胞中的先于B而存在信息(具有位置信息的情况便是如此),还是A细胞可以从其环境中获取信息。在后一种情况下,A和B之间的指导关系相当于在稳态电路中发现的常见的传感器-效应器关系:传感器细胞监测一个稳态变量的值,并产生信号,指导效应器细胞在期望的方向改变该值。
3.4 提供者-消费者关系
这一类细胞关系是基于细胞之间的存在依赖关系,其中细胞A依赖细胞B在特定的组织龛中存在。类似于生态系统中生物之间的营养关系,一个细胞可能依赖另一个细胞提供的资源来生存。这些资源可以是谱系限制的GFs、代谢物以及供应者细胞提供的其他信号,这些信号对于需要它们的细胞的生存至关重要。另一种存在依赖性是当一种细胞类型调节组织微环境使其允许另一种细胞类型存在时,如ECM特性如何影响细胞的粘附和存活。与信息不对称所定义的传感器-效应器关系不同,这种关系是由资源不对称所定义的 (图4d)。
3.5 层级关系
类似于生态系统,组织细胞往往表现出不同细胞类型之间的等级关系,其中某些细胞类型比其他细胞类型对组织的完整性、功能和组成更重要。例如,虽然组织驻留T细胞为其所在的组织增加了一些功能,但它们并不是组织所必需的。事实上,通过基因或其他手段消除这些细胞不会破坏组织结构。可以用一个类似于图4e中马斯洛的人类需求层次的重要性金字塔来说明这一点。细胞的层次性还体现在特定细胞作为组织成分的调节者的作用。这类似于生态系统中的关键物种。关键物种不成比例地影响着它们所在的生态系统。尽管关键种的质量或数量可能相对较小,但关键种的存在对生态系统中物种的组成和比例有很大的影响。
3.6 相互排斥关系
虑到不同组织的细胞组成,我们经常会看到一些细胞类型是相互排斥的,并不共存于同一组织隔室中 (图4f)。相互排斥可以发生在几个原因中:一种细胞类型的存在可能使组织环境不允许另一种细胞类型存在,两种细胞可能竞争相同的资源或功能不兼容,或者两种细胞可能由于缺乏任何发育或生理情况而被排除在同一生态位中。
4. 组织的模块化构成:
最小的组织单元和更高阶的组织模块
4. 组织的模块化构成:
最小的组织单元和更高阶的组织模块
模块化是生物系统的一个共同特征。模块化描述了一个由可分解的组件或模块组成的复杂系统,使得每个模块由相互作用的元素组成,而不是与模块外的元素相互作用。尽管模块化在细胞和分子水平上得到了很好的理解,但组织模块化并没有得到很好的定义。这里从三个角度来考虑组织中的模块:组织组成、结构和功能的模块。
4.1 组织模块的组成
基本的组织成分来源于原始的上皮-间充质组织单元。在所有的后生动物中,组织中的细胞类型被分为上皮细胞和间充质细胞,上皮-间充质相互作用是许多基本的发育过程。鉴于其古老的起源和在组织模式化中的基础作用,上皮-间充质模块可以被认为是组织细胞的一个基本单元。上皮和间充质细胞之间的相互作用由FGF,BMP,hedgehog (Hh),Wnt和Notch家族成员介导,它们控制上皮和间充质分化和形态发生。上皮-间充质模块既说明了主要支持的功能关系,也说明了指导性的关系。
在复杂的后生动物中,包括脊椎动物,上皮-间充质单元作为主要的支持关系被扩大。其中间充质细胞进一步分化为中胚层细胞类型,具有更特殊的支持功能,这几乎是脊椎动物组织的普遍特征,它们构成了一个最小的组织单元:基质细胞,毛细血管内皮细胞和组织驻留的巨噬细胞 (图5a)。基质细胞主要由成纤维细胞组成,负责结构支撑。组织定居的巨噬细胞存在于许多器官特异性的品种中,包括小胶质细胞 (大脑),Kupffer细胞 (肝脏),破骨细胞 (骨),肺泡巨噬细胞 (肺)。与巨噬细胞一样,基质和微血管内皮细胞也具有组织特异性特征,这可能反映了其主要功能细胞的多样性。大多数脊椎动物组织中含有额外的支持细胞类型,包括组织驻留淋巴细胞,树突状细胞,以及一些组织中的脂肪细胞和肥大细胞。这些代表了在组织稳态和防御中专门用于特定支持功能的细胞。
4.2 组织模块的结构
最基本的结构组织模块由两层上皮和间充质细胞以及间充质细胞产生的基质组成。为了形成这种结构,上皮细胞受细胞-细胞相互作用的控制,从而形成上皮层,间充质细胞受细胞-基质相互作用的控制。大多数上皮组织可以追溯到原始上皮-间充质-基质的设计。其他类型的组织也呈现出这种简单设计。例如,在骨骼肌中,肌腱将肌肉附着在骨上,并作为一种类似于间充质-基质设计的基质。上皮组织中的高级空间排列,如组织极性和分支形态发生,是上皮细胞和间充质细胞之间通过诱导信号相互作用而产生的模式。
虽然哺乳动物组织是简单的上皮-间充质-基质设计的阐述,但在某些情况下,与原始设计的关系可能会被形态发生的特殊特征所掩盖。这些案例体现了连续变换(一个来自数学和拓扑学的概念)。在连续变换中,物体的形状可以被修改,使得物体的部分可以急剧地膨胀/收缩。因此,物体上点与点之间的相对距离会发生变化,从而导致整体形状的剧烈变化。与这一概念类似,某些组织并没有显示出可识别的结构模块,因为它们是通过扭曲原始图案的基本组织设计的连续变换而进化出来的。这种连续的转变可以在大脑和内胚层来源的器官的结构中看到。定义结构模块的一个重要特征是形成边界。组织中边界的形成是一个自组织的过程,最终导致组织的区室化。
4.3 组织模块的功能
功能单位是由若干个细胞类型来定义的,这些细胞类型将功能进行划分,以集体执行一个联合的功能。这种分工可以在从互补关系到执行-支持关系的连续谱上找到。基于互补关系的功能单位实例包括运动神经元与骨骼肌、周细胞与内皮细胞、破骨细胞与成骨细胞 (图5c)。某些功能单位可以在不同的组织中重复,而这些功能模块中的细胞类型的身份可能会有所不同。例如,ECM的组成通常主要由成纤维细胞和巨噬细胞调节,但在大脑中,ECM也由星形胶质细胞产生。在组织水平上,基本的功能单位可以组合成更高级的单位,如肺泡、小肠绒毛、肾单位等。高阶功能单元存在多个拷贝,并且经常围绕微血管组织组成。
5. 自组织和涌现
5. 自组织和涌现
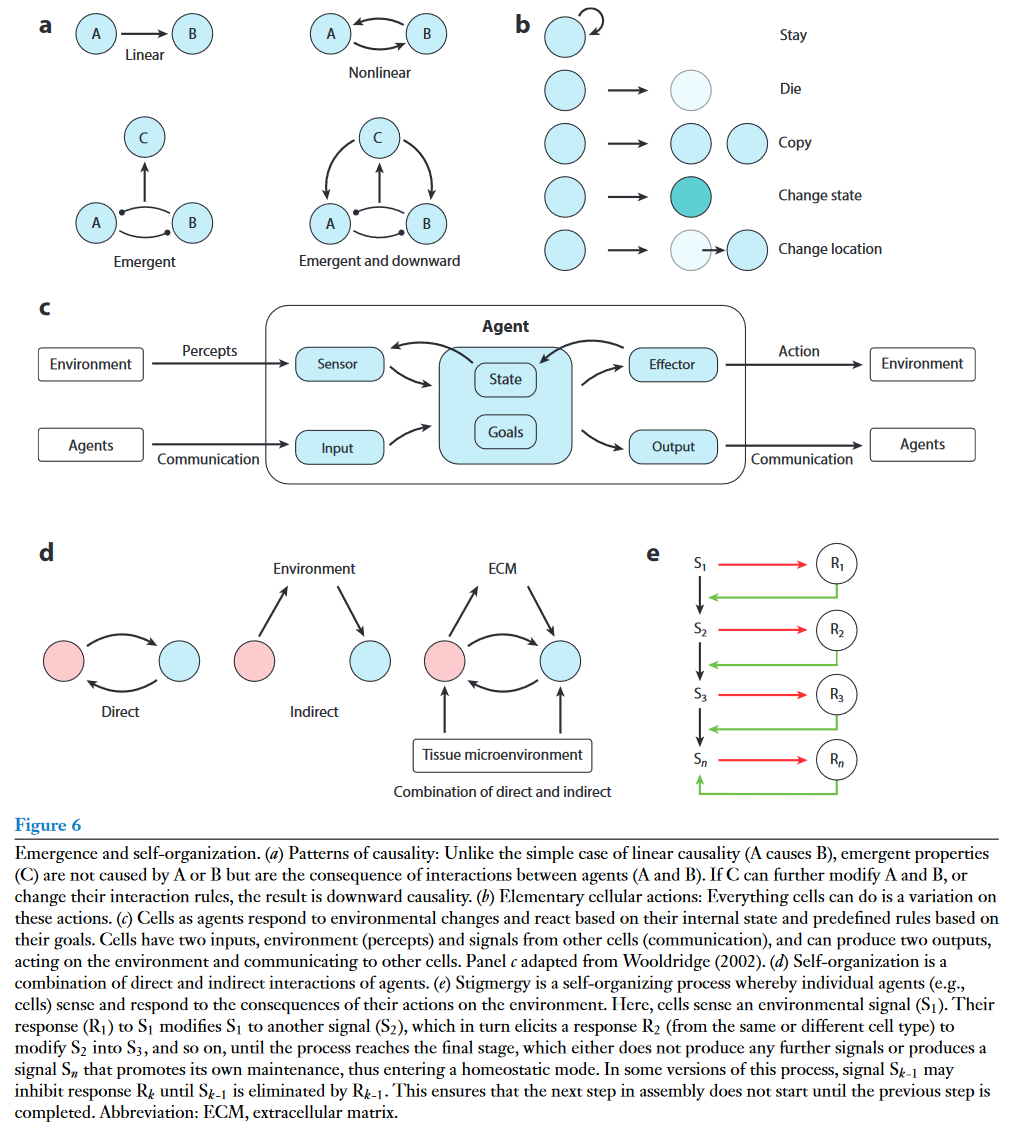
上面讨论的许多组织特征,包括空间模式、组成和功能,都是新出现的特性,因此它们不能根据组织中涉及的单个细胞、基因或信号通路的详细知识来解释。涌现属性是指系统中具有足够多的多样性成分(或能动者),这些成分(或能动者)可以按照某种预先指定的规则相互作用的产物。因此,为了理解一个复杂系统,人们需要定义其产生涌现性质的组件之间的相互作用规则 (图6a)。
社会性昆虫最能体现昆虫的涌现性和自组织性:白蚁可以构建复杂的结构,但个体昆虫完全不知道它们共同产生的最终产物。社会性昆虫的例子突出了基于分散控制的自组织和涌现的一般原理:( a )个体对局部可用信息做出反应;( b )这些信息通过微环境的改变直接或间接地编码在来自其他因素的信号中;和( c )根据一些预定义的规则来响应这些信号。重要的是,即使是少量的简单规则也足以产生复杂的模式和行为——系统的涌现属性。对于组织来说,个体则是细胞,它们根据一组预定义的、细胞类型特异性的规则相互作用并对其环境作出反应。指导细胞相互作用的预定义规则基于细胞的目标。例如,作为目标的内稳态导致反馈相互作用,将组织变量维持在固定水平。基于相互作用规则、细胞内部状态和外部环境条件,细胞可以采取几种可能的行为之一:它可以停留在相同的细胞状态,死亡,增殖,或改变其状态或位置 (图6b , c)。
细胞相互作用的规则大概属于几个程序,每个程序都是兼容规则的集合。一些程序被用于核心发育过程,如组织极化、空间图案化、管腔形成、分支形态发生和隔室边界形成等。少数家族的信号 (BMP、FGF、Hh、Wnt、Eph、Notch) 在其中起着突出的作用。此外,机械力在组织发育和图案化中也发挥的重要作用。一旦发育程序建立了基本的组织框架,细胞就会转换到控制组织组成和稳态的程序中。在组织修复和肿瘤发生过程中,发育程序可以重新激活,并可能伴随着短暂地关闭稳态程序,因为它们可能与组织修复不相容。例如,在女性生殖器官中会发生周期性的组织重塑,而诱导性组织重塑通常由炎症信号协调。
6. 细胞间通讯规则
6. 细胞间通讯规则
6.1 功能需求和生长因子生产的控制
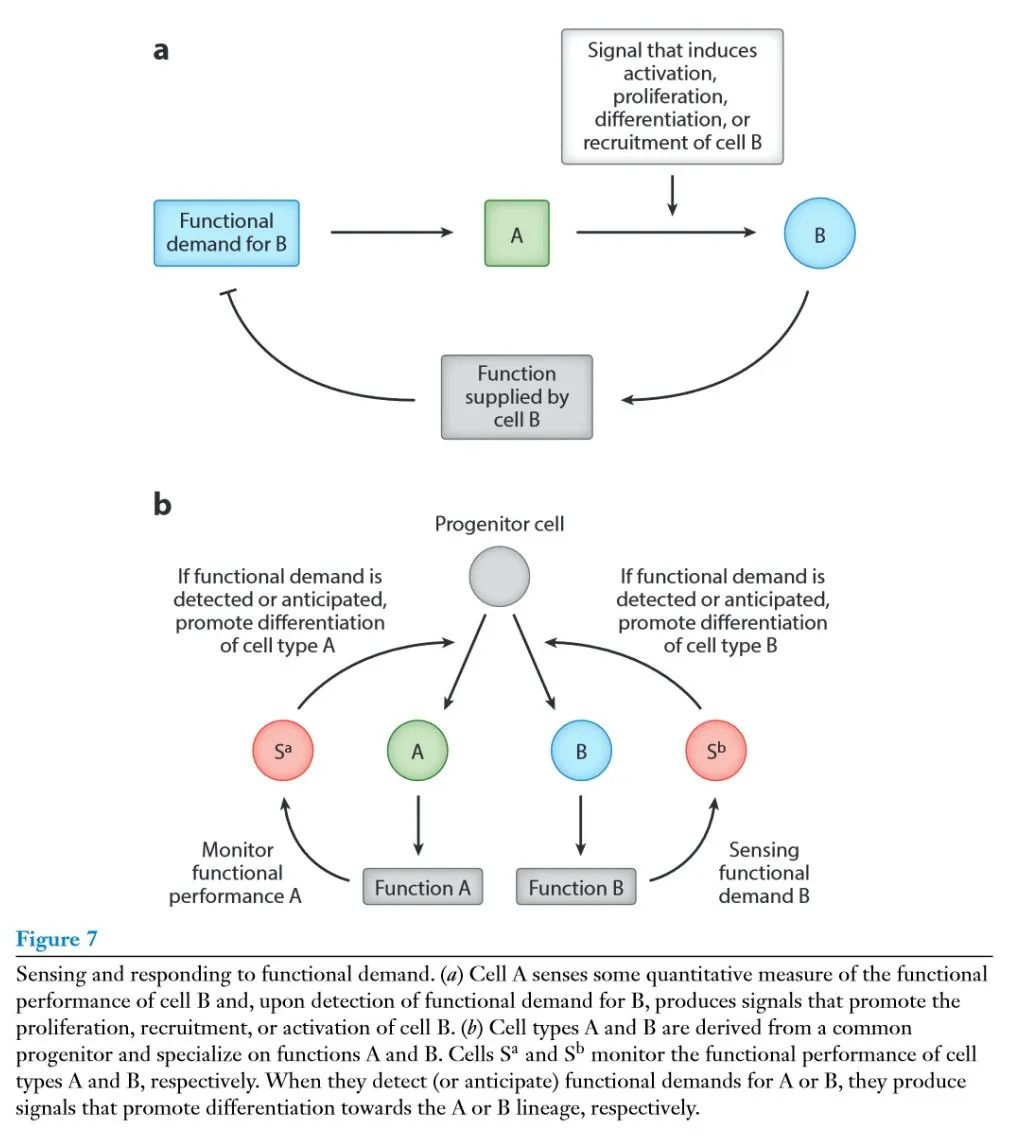
组织的细胞组成由特定谱系的生长因子(growth factors,GFs)的局部可用性决定,这些GFs是特定细胞谱系生存、增殖和分化所必需的,例如,巨噬细胞的M – CSF,成纤维细胞的PDGF。控制GF产生的逻辑在很大程度上是未知的:组织内的单个细胞如何决定在任何给定的时间内产生哪种GF,以及多少,以确保组织内细胞的正确细胞组成和空间排列? 实现这一目标的一种方法是通过设计细胞交互电路来维持GF交换中所涉及的细胞类型的稳定比例。例如局部组织缺氧表示对氧的需求,由血液内皮细胞供应。转录因子HIF – 1a对缺氧的感知导致VEGF的表达,进而促进血管生成。这个例子提出了一个调控谱系特异性GF产生的一般原则:如果细胞类型A是控制细胞类型B的增殖、存活、激活、分化或招募的信号S (Sa) 的供应者,那么细胞A产生Sa是通过感知细胞B执行功能的需求来控制的 (图7a)。通过这种方式,给定组织腔室中B细胞的数量或活性被自动调整以适应需求。
从功能需求的角度来看,有以下几点启示:第一,应该有有限的功能需求集合,有的是普适性的 (例如,氧气输送),有的是组织特异性的 (例如,感知肠细胞的吸收功能)。其次,对于每一种功能需求都必须有相应的感知机制。第三,每个功能需求都应该有对应的专用信号 (如M – CSF对应于巨噬细胞中的需求) 。当一个以上的信号对应一个给定的功能需求时,它们的生产很可能是协同调控的。除了细胞之间以及细胞与微环境之间的相互作用,细胞外基质也是组织局部信号的另一个重要来源。
6.2 细胞外基质和共识主动性:构建和解释细胞环境
5) ECM是由基质金属蛋白酶 (MMPs) 介导的有限蛋白水解释放的可扩散信号 ( matricryptins) 的来源。
ECM在介导组织背景中细胞间的间接相互作用中具有重要作用 (图6d)。这种通过环境的交流产生了一种独特的自组织形式,被称为共识主动性(stigmergy),最初是为了解释社会昆虫在筑巢过程中的集体行为而提出的。ECM的复杂结构是ECM成分逐步成熟、沉积、组装和分解的产物。细胞可以检测ECM构建的中间状态产生的信号,并响应进一步修改中间状态,直到达到最终的完全组装状态。此时,沉积或修饰ECM的信号将被消除,系统将达到稳态,即内稳态 (图6e)。当ECM发生损伤时,中间信号再次被揭示,触发同样的一组递归动作来修复损伤。组织内不同的细胞类型可以执行不同的规则,这进一步耦合了ECM结构和细胞组成。
总结
总结
最近在单细胞基因组学、离体类器官和成像方面的进展为组织生物学提供了重要的数据和见解。随着类器官技术的不断成熟,我们应该很好地解决组织自组织的基本问题。然而,目前类器官的局限性在于缺乏对ECM组成的实验控制,所需的GFs是外部供应给系统的,而不是由细胞按照自己的逻辑产生。随着这一障碍的克服,应该可以系统地探索GF产生的规律和产生组织结构涌现特性的其他细胞通讯形式。本文提出了一些新的观点,以帮助发展组织生物学的概念框架。复杂系统理论所揭示的原理对于正确理解组织作为执行细胞相互作用的简单规则的产物是特别相关和必要的。本文认为,定义这些规则的完整目录将产生一个组织生物学的现代范式,并将帮助理解和最终治疗相关的疾病。
参考文献:
|通讯作者

Ruslan Medzhitov,美国国家科学院院士、耶鲁大学医学院免疫生物学教授、Tananbaum人类生物学理论与分析中心研究员、霍华德休斯医学研究所研究员。
生命复杂性读书会招募中
在生物学中心法则的起点,基因作为生命复杂系统的遗传信息载体,在生命周期内稳定存在;而位于中心法则末端的蛋白质,其组织构成和时空变化的复杂性呈指数式增长。随着分子生物学数十年来的突飞猛进,尤其是生命组学(基因组学、转录组学、蛋白质组学和代谢组学等的集合)等领域的日新月异,当代生命科学临近爆发的边缘。如此海量的数据如何帮助我们揭示宇宙中最复杂的物质系统——“人体”的构成原理和设计原理?阐释人类发育、衰老和重大疾病的发生机制?
集智俱乐部联合西湖大学理学院及交叉科学中心讲席教授汤雷翰,国家蛋白质科学中心(北京)副研究员常乘、李杨,香港浸会大学助理教授唐乾元,北京大学前沿交叉学科研究院研究员林一瀚,中国科学院分子细胞科学卓越创新中心博士后唐诗婕,共同发起「生命复杂性:生命复杂系统的构成原理」读书会,从微观细胞尺度、介观组织器官尺度到宏观人体尺度,梳理生命科学领域中的重要问题及重要数据,由生物学家提问,希望促进统计物理、机器学习方法研究者和生命科学研究者之间的深度交流,建立跨学科合作关系,激发新的研究思路和合作项目。读书会从2024年8月6日开始,每周二晚19:00-21:00进行,持续时间预计10-12周。欢迎对这个生命科学、物理学、计算机科学、复杂系统科学深度交叉的前沿领域感兴趣的朋友加入!
推荐阅读
微信扫一扫,分享到朋友圈











