理论生物学家和复杂系统科学家 Stuart Kauffman 与合作者最近发表文章,提出一种新的生命定义,根据这个定义,生命在宇宙中的出现是意料之中的,而其不断创造性的开放式进化则无规律可循。生命有机体是实现了催化闭合、约束闭合和空间闭合的康德式整体。本文将集体自催化集(Collectively Autocatalytic Sets)和相邻可能理论(Theory of the Adjacent Possible)这两个数学理论结合在一起,指出生命可以看作是宇宙演化过程中的一个相变过程。本文是对这篇文章的翻译。
关键词:相变,催化闭合,集体自催化集,约束闭合,康德式整体,牛顿范式,通用构造器
![]()
Stuart Kauffman, Andrea Roll | 作者
John | 译者
小木球 | 审校
梁金 | 编辑
Is the Emergence of Life an Expected Phase Transition in the Evolving Universe?
https://arxiv.org/abs/2401.09514v1
摘要
引子
1. 对生命的定义
2. 第一个奇迹:生命的涌现是可预期的相变——TAP理论和RAF理论。
3. 第二个奇迹:生物圈的演化是一种传播性的、不可推导式的构造,而不是必然的演绎。不存在定律。演化是不断创造的过程。
4. 新的观察和实验:宇宙中存在其他生命吗?
总结
我们提出了一种新的生命定义,根据这个定义,生命在宇宙中的出现是意料之中的,而其不断创造性的开放式进化则是无规律可循的。生命有机体是实现了催化闭合、约束闭合和空间闭合的康德式整体。
本文中,我们首次将两个成熟的数学理论,即集体自催化集(Collectively Autocatalytic Sets)和相邻可能理论(Theory of the Adjacent Possible)结合在一起。前者认为,在宇宙的化学演化过程中,随着分子的多样性和复杂性不断增加,必然会出现以分子繁殖(复制)为特征的一阶相变;后者认为,在宽松的假设条件下,如果系统开始时只有少量初始分子,每个分子都可以与自身或其他分子的复制品结合生成新的分子,随着时间的推移,分子种类的数量会缓慢增加,但随后会出现双曲线式的爆炸式增长。
将这两种理论融合到一起则提示,生命可以看作是宇宙演化过程中的一个相变过程。我们熟悉的软件和硬件之间的区别用于讨论活细胞时,就失去了意义。因此,在本文中,我们提出了研究新陈代谢系统发育的新方法、在系外行星上寻找生命的新天文方法、寻求最初级生命出现的新实验,以及通向具有模板复制和编码功能的原核生物的可检验的连贯途径的蛛丝马迹。
埃尔温·薛定谔在 1944 年出版的名著《生命是什么》中,精辟地提出了非周期性晶体是生命体秩序的源泉(1)。但他没有回答这个问题。八十年后,在前人研究的基础上,我们相信对此可以勾勒出一幅连贯的图景。宇宙中可能有一百万亿颗宜居行星。宇宙中生命的出现是一个奇迹,但在我们看来,这是一个意料之中的奇迹。一旦生命出现,它的演化就具有根本性的创造力,而且不能仅仅以物理学为基础。强还原论在这里显示了它的局限性。生命是一个双重奇迹。
目前学术界对生命并没有统一的定义。在本文中,我们希望就生命的定义问题构建如下描述:生命是一个非平衡、自我复制的化学反应系统,它能够实现:1) 集体自催化,2) 约束闭合,3)空间闭合;4)由于以上三点,生命是康德式整体。我们将在下面解释这些概念。
1.1 集体自催化集
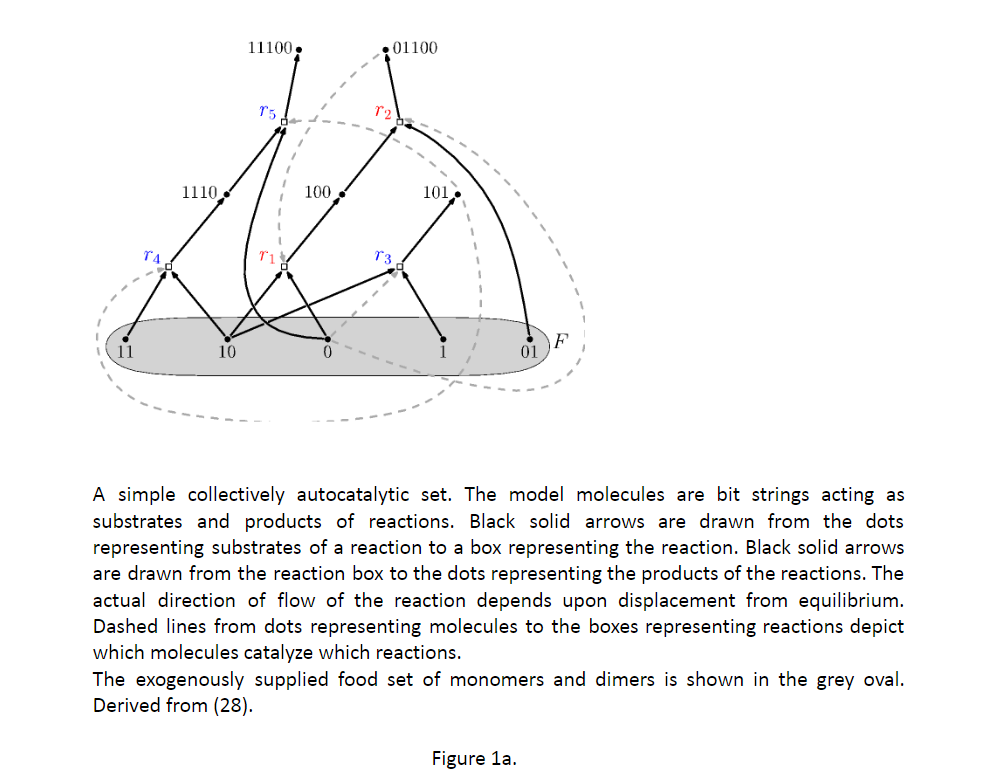
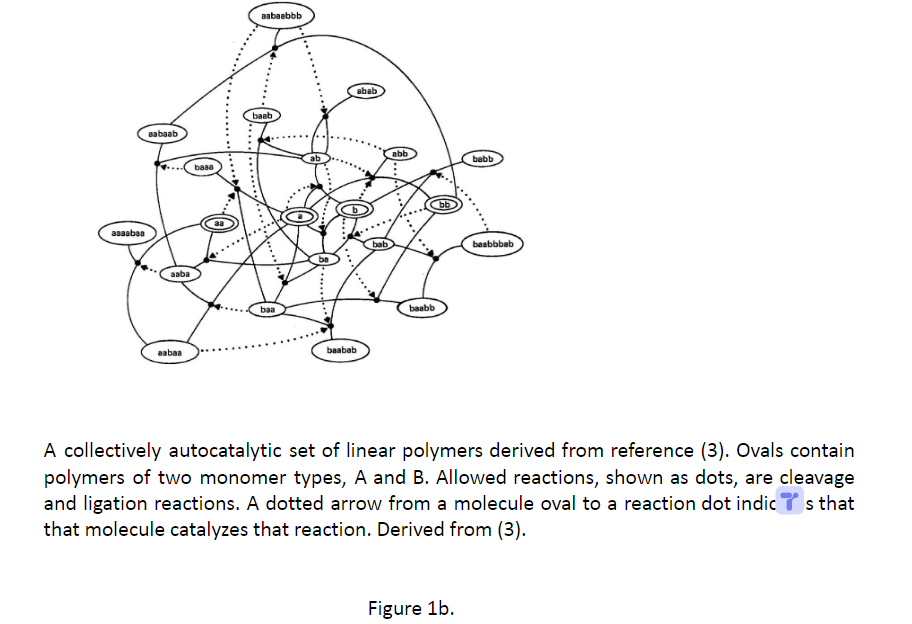
集体自催化集(CAS)是一个开放的化学反应系统,由外源分子和能量结构单元供给,具有以下特性:形成该集合中每个分子的最后化学反应步骤,由该集合中的至少一个分子催化,或由食物集中一个分子催化。图 1a 显示了一个简单的示例 (2)。图 1b 显示了一个更复杂的示例 (3)。
生命必须基于“由可进行模板复制的多核苷酸构成”的概念,主导了生命起源领域约 50 年 (4,5)。然而,基于“裸的可复制的 RNA”的复制至今尚未实现(6)。尽管如此,这个目标还是有可能实现的。
图1a 一个简易的集体自催化集。模型分子是充当底物和反应产物的字符串。黑色实心箭头从代表反应底物的点画到代表反应的方框。黑色实心箭头从代表反应的框指向代表反应产物的点。反应的实际流动方向取决于平衡位移。从代表分子的点到代表反应的框的虚线描绘了哪些分子催化哪些反应。外源提供的单体和二聚体食物组显示在灰色椭圆形中。源自(28)。
图1b 源自参考文献 (3) 的线性聚合物的集体自催化集。椭圆形包含两种单体类型 A 和 B 的聚合物。允许的反应(显示为点)是裂解和连接反应。从分子椭圆形到反应点的虚线箭头表示该分子催化该反应。源自(3)。
“可进行基于模板的复制的双链 RNA 序列”这一熟悉的概念是集体自催化集的一个具体例子。每条链都是合成另一条链的模板催化剂。然而,集体自催化的概念要广泛得多。
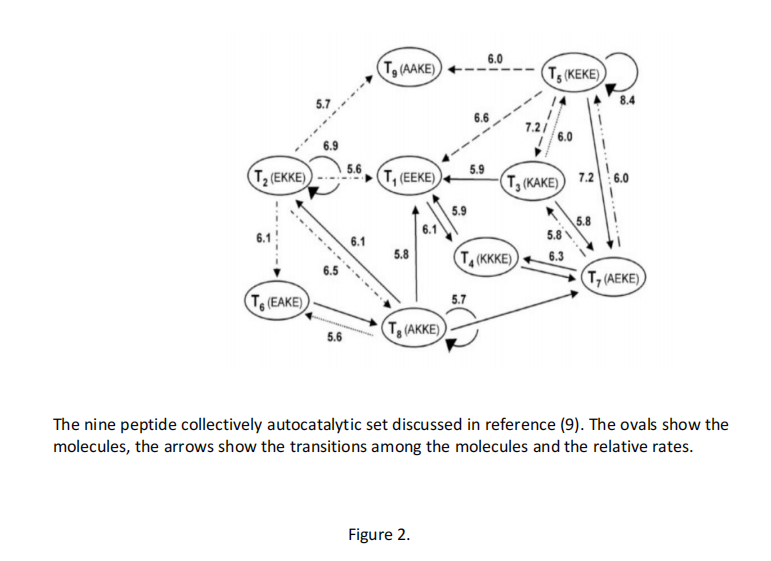
与获得可复制的 RNA 序列的希望形成鲜明对比的是,DNA、RNA 和肽的集体自催化集已经被构建出来。第一个是 DNA 集体自催化集,由 G. von Kiedrowski 构建 (7)。N. Lehman 及其同事实现了 RNA 集体自催化集 (8)。该集合可以根据其构建块自发地自我组织。图2显示了由 G. Ashkenasy 构建的九种肽的集体自催化集 (9)。还考虑了脂质的自催化集 (10)。
图2 参考文献(9)中讨论了九种肽的自催化集合。椭圆形显示分子,箭头显示分子之间的转变和相对速率。
这些结果具有根本意义上的重要性。它实现了创建自我复制的开放的化学反应系统。
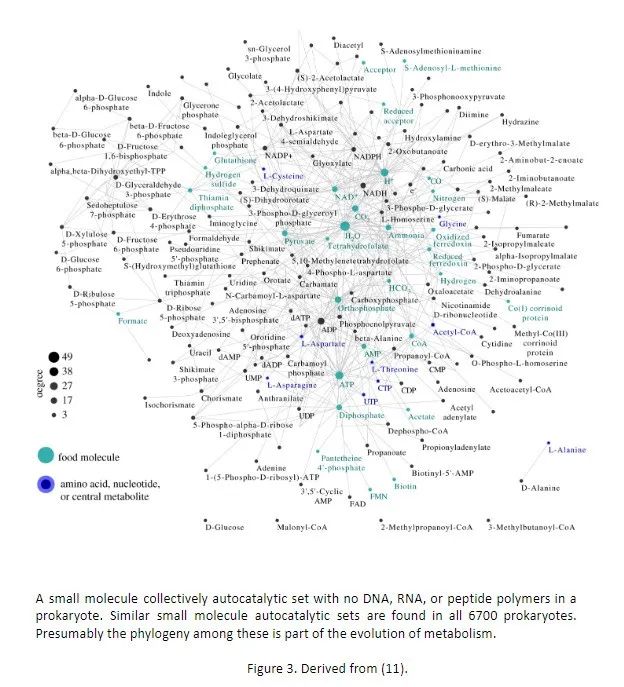
目前令人震惊的证据表明,在所有 6700 种原核生物中都存在不含 DNA、RNA 或肽聚合物的小分子集体自催化集,图 3 (11,12)。这些小分子自复制集合包含数百到数千个小分子以及它们之间的反应。这些自催化装置可以合成多种氨基酸和 ATP,是通过计算机模拟来识别的。它们能否实现体外(化学水平的,译注)繁殖仍有待证明。
在这所有 6700 种原核生物中存在着小分子集体自催化集的证据强烈表明,宇宙中第一个能够自我复制的化学系统可能正是这样的反应集合。我们在下面将证明,此类集合的出现是可预期的。
图3. 由文献 (11) 导出。原核生物中存在不含 DNA、RNA 或肽聚合物的小分子集体自催化集。在所有 6700 种原核生物中都发现了类似的小分子自催化集。据推测,它们之间的系统发育是新陈代谢进化的一部分。
小分子自催化集的鉴定与关于生命起源中模板复制多核苷酸必要性的持续争论有关。这样的“裸 RNA 基因”必须进化出 RNA 序列来催化相关的新陈代谢,从而创建和维持自己的构建模块。然而,完全没有理由解释为什么这种相互关联的新陈代谢本身也具有集体自催化作用。这种考虑增加了人们对分子复制起源是通过小分子集体自催化集的出现的信心。
1.2 生命:康德式整体、催化闭合、约束闭合、空间闭合
1790 年代,哲学家伊曼纽尔·康德(Immanuel Kant)引入了一个基本概念:一个有机的存在物具有部分为整体而存在,并通过整体而存在的属性 (13)。康德的洞察力已经沉睡了 230 年。所有生命体都是康德式整体,为其部分而存在并通过其部分而存在。你也是一个康德式整体。你通过你的身体部位而存在——心、肝、肾、肺、脑。你的部分是通过你,做为整体而存在的。你繁衍后代,你的孩子继承你的部分。
所有生命体都是康德式整体。这包括分类尚未定论的的病毒。在细胞环境内,病毒是可繁殖的康德式整体。做为细胞的一部分,病毒的各个部件会产生病毒这些部件的多个副本,这些副本会自组装成成熟的病毒整体。有趣的是,康德式整体做为是否是生命的定义,将病毒判定为活的生命。
康德式整体是一类特殊的物理动力系统。晶体不是康德式整体,晶体的原子可以不作为晶体的一部分而存在。砖头也不是康德式整体,而细胞显然是康德式整体。
集体自催化集(如图 2中的9肽组)可实现催化闭合。系统中的每个反应均由系统中的至少一个分子催化。所有活细胞都实现了催化闭合。活细胞中没有分子催化其自身的形成。但在细胞繁殖(复制)时,活细胞中的一组分子(做为一个整体)实现了催化闭合(14,15,16)。
实现催化闭合的系统同时也是康德式整体。图2中9个肽集体自催化集中的每个肽都是整体的一个部分,每个肽都存在于九个肽整体当中,并通过这九个肽成为整体,这九个肽的相互催化使所有部分能够成为一个不可分割的整体。
活细胞,包括在所有 6700 种原核生物中发现的小分子集体自催化集,实现了新近公认的深刻特性:约束闭合 (Constraint Closure)(17)。其热力学功是将能量限制释放到几个自由度中(18)。一个例子是底部装有火药的大炮,火药旁边有一个炮弹。当能量爆炸时,大炮既是边界条件又是约束,限制能量的释放以将炮弹从炮膛中炸开。热力学工作是在炮弹上完成的。因此,在非平衡过程中,如果不存在这3个能量释放约束,则无法进行热力学工作(19)。
牛顿没有告诉我们边界条件从何而来。例子中的大炮就是边界条件。但大炮从哪里来呢?关键的答案是组装大炮需要热力学做功。我们可以得出结论:没有约束,就没有做功。但构建相关约束通常需要做功。因此:没有约束,就没有做功;同样,没有做功,就无法构建和描述约束条件。如何刻画这个做功-约束循环是一个新课题(19)。
Maël Montévil 和 Matteo Mossio 在 2015 年首次定义了约束闭合(Constraint Closure),(17):考虑一个具有三个非平衡过程 1、2 和 3 的系统。考虑三个约束 A、B 和 C。让 A 约束过程 1 中的能量释放来构造 B。让 B 约束过程 2 中的能量释放来构造 C。让 C 约束过程 3 中的能量释放来构造 A(见图 1a、1b 和 2)。
上述系统实现了一个显著的特性:约束闭合(闭包)。一组约束条件(此处为 A、B 和 C)将一组过程(此处为 1、2 和 3)的能量释放限制为几个自由度,因此热力学功构造了完全相同的一组约束, A、B 和 C!该系统实际上是通过构建自己的边界条件约束来构建自身的热力学做功,该边界条件约束构建相同的边界条件的能量释放。
约束闭合是一个全新的概念,以制造汽车为例。汽车是由一些零件组合出精确装置,包含通过相互撞击来释放能量的部件。并完成汽油燃烧,活塞活动,车轮转动等一系列功能。但(与活细胞不同的是),汽车并没有能力构建自身对于能量释放的边界约束条件。
所有自催化分子反应系统都实现了催化闭合和约束闭合。这些都是康德式整体。例如,在图 2 中的 9 肽集体自催化集中,每个肽通过结合下一个肽的两个片段来充当连接酶。通过定向两个片段,作为连接酶的肽降低了两个片段连接的激活势垒,以产生下一个肽的第二个拷贝。当肽键形成时,进行热力学工作以构建下一个肽。因为这个集体自催化肽系统中的所有反应都是如此,所以该系统作为一个整体实现了催化闭合和约束闭合。该系统自行构建。而且这个系统也是一个康德式整体。
最重要的是所有活细胞都实现约束闭合。细胞构建了能量释放的边界条件,能量释放也构建了同样的部件构成的边界条件。细胞可以自行构建,而计算机和汽车却不能构建自身。
可以自我繁殖(自我复制)的细胞从根本上来说并不是冯·诺依曼的自我复制自动机(20),后者基于“通用构造器”运转。为了构造任何特定的东西,通用构造器需要特定的“指令”。指令以物理方式编码并内置于通用构造器内部。以物理形式体现的指令发挥着双重作用:它们用于构造通用构造器的副本,在该副本中构造物理指令的物理副本,然后将其插入。物理指令的双重作用恰恰构成了软件和硬件之间的区别。与此形成鲜明对比的是,活细胞通过催化和约束闭合,特异性地构建自身。活细胞并不是需要额外指令集才能运行的通用构造器。例如,图 2 中可自我复制的 9 肽组就没有一套额外(可单独分离的)“指令”来编码其如何形成。这里软件和硬件的概念区分是不适用的。
Paul Davies (21) 指出,在活细胞的背景下,基因以及转录和翻译装置实际上是所有可能编码的多肽的通用构造器(而非只是这9肽组合,译注)。基因可以被视为一组指令集。然而,基因所在的活细胞本身并不是通用构造器。它只专门地构建自己(译注:原文含义是活细胞不具备通用性,然而译者认为此处并不准确。因为活细胞的复制翻译等制造系统是有一定通用性的,这也是合成生物学的基础)。如果它的数千个基因中的每一个都被编码某种随机多肽的随机DNA序列取代,那么合成这些新蛋白质的细胞几乎肯定会灭亡。
生命体已经进化形成嵌套的康德式整体。原核生物是一阶康德式整体。真核细胞是与线粒体和叶绿体的共生体,(22, 23),是包含一阶康德式整体的二阶康德式整体。多细胞生命体是包含二阶和一阶康德式整体的三阶康德式整体。
2. 第一个奇迹:生命的涌现
是可预期的相变——TAP理论和RAF理论
在第二部分中,我们将表明,随着系统中分子种类多样性的增加,集体自催化集的出现是化学反应网络中可预期(必然发生)的相变,并且分子之间的反应多样性将增加得更快。我们首先讨论在足够丰富的化学反应网络中集体自催化(RAF)的相变。然后我们将 RAF 理论与 TAP 理论结合起来,得出化学物质的多样性不断增加,其中 RAF 相变最终必须发生。这个“TAP-RAF”理论是本文最新的发现。
生命分子是由 CHNOPS(即碳、氢、氮、氧、磷、硫)等多种元素组成的整体(24)。一个分子由例如 10 个原子组成,每个原子与一个或两个其他原子键合,构成不同的多样性模块并组成一个个分子,这些多样性模块的集合同时也是化学反应网络中反应底物的集合,反应底物组分的集合构成了细胞化学反应网络本身。
举一个简单的例子,如有两个结构单元 A 和 B 组成的线性聚合物(ABBABAABBA),该聚合物有十个结构单元。通过破坏相邻构建块之间的 9 个键中的任何一个即可轻松看出,这种单一聚合物可以通过 9 种不同的方式构建,随着系统中分子组合复杂性的增加,整个反应系统中反应 R 与分子 M 的比率 R/M 增加。
化学反应图是一个“二分图”,由代表分子种类的点和代表分子相互转化的不同反应的方框组成。箭头从代表反应底物的点指向反应框。箭头从反应框指向反应产物。该图只表示化学反应网络的结构,而不是由化学平衡决定的化学反应热力学流动方向(3, 15)。
下一步很关键:如果我们知道哪些分子催化了哪些反应,我们可以用从相关催化剂分子到它催化的反应的虚线箭头来表示。这种结构称为二部超图,见图 1b。给定一个特定的二部超图,可以检查它是否包含一个集体自催化集。图 1b 中的反应系统是此类二部超图的一个示例,其中包含集体自催化集 (3)。
一般来说,我们不知道哪个分子催化哪个反应。在缺乏特定知识的情况下,理论和见解可以通过单纯的假设来进行,即每个分子都有一定的概率 Pcat 来催化每个反应。这个简单的假设已经带来了显著的结果。随着分子及其反应多样性的增加,集体自催化集作为一级相变而出现。接下来我们看看为什么 (3, 15, 25)。
1959 年,两位数学家 Erdos 和 Renyi 发表了一篇关于随机图性质的开创性论文 (26)。一个随机图包含一组单点、节点或顶点。每个点可以通过线连接到一个其他点、或连接到一定数量的其他点,或不连接任何点。
Erdos 和 Renyi 提出了一个精彩的问题:从 N 个节点开始。随机选取一对节点并用线连接它们。迭代地继续选取随机的节点对并用线连接它们。令 N 为节点数。在这个过程中的任何一步,让连接节点的线的数量为 L。考虑比率:L/N。对于固定 N,当 L 增加时,图会发生什么?
当 L/N 的比率增加到 0.5 时,奇迹就会发生。突然间,一个巨大的连接组件或网络出现了。在这样的巨型组件中,每个节点都直接或间接连接到该巨型组件中的所有其他节点。比率 L/N = 0.5 是一阶相变。给定固定数量的节点 N,随着 L 增加并超过 L/N = 0.5,几乎肯定会出现一个巨大的连通分量。
集体自催化集(也称为 RAF)的涌现是由于二分化学反应超图中的相变的发生是同样的。考虑给定的二分化学反应图,当我们增加表示催化任意分子的任意给定反应概率 Pcat。根据哪些分子催化哪些反应随机分配Pcat的取值。该系统是否包含集体自催化集?当 Pcat 值为某些特定取值时,许多反应被催化,以至于它们形成了一个巨大的集合,即集体自催化集(3,15,26)。这正是在足够丰富的非平衡化学反应体系中发生的可产生分子复制功能(现象)的一级相变。
更重要的是,当保持 Pcat 恒定,并增加系统中分子的数量和元素复杂性。反应与分子的比率 R/M 必然增加。当系统中的分子具有一定的复杂性,及 R/M 比率时,一个集体自催化集(RAF)就必然出现(概率接近 1.0)(3)。这是基本的一级相变,通过它可以产生自我复制的分子系统,例如 6700 种原核生物中都包含的小分子集体自催化集。
本文作者中的一位同事 (14) 在 1971 年创建了第一个模型,展示了集体自催化集的自发出现。1986 年发表的两篇文章,其中一篇由Kauffman单独发表,其中的定理证明了集体自催化集的出现是可预期的(必然的) (15)。第二个由Farmer, Packard 和 Kauffman 共同提出,该研究实现了基于“二元聚合物模型”的计算机代码(3)。每个模型中的聚合物均由两种单体 A 和 B 组成,聚合物的最大长度 L 是固定的,所有 2L+1 种可能的聚合物都可以成为系统的一部分。该反应系统由含可能聚合物子集组成的“食物集”维持。该模型只允许裂解和缩合反应,每种聚合物具有相同且固定催化概率的 Pcat 来催化每种反应。
结果是令人信服的:对于固定的 Pcat,随着允许的最长聚合物的长度 L 增加,潜在聚合物的多样性随着 R/M 比率的增加而增加,总体自催化集出现的概率接近 1.0。图 1b 就是这样一套集体自催化装置。
接下来几十年的大量工作(27,28,29)已经证明:一、随机分配哪种聚合物催化哪种反应的规则可以是均匀的或幂律的,这二者之间并没什么区别。二、包含序列识别的模型几乎没有什么区别。三、每种聚合物必须催化的反应数量在 1 到 2 之间。以上在化学上却是合理的。四、重要的是,Hordijk 和 Steel 发现这种集体自催化集包含“不可约自催化集”,它们共同形成包含许多此类不可约组的更复杂的系统 (28, 29)。Vassas 等人指出,这种不可化约的自催化集,每个都能够充当独立的复制子,可以充当可以继承并分配给子集的“基因”。因此,这种没有“基因组”的简单系统可以进化(29)。这种不可约自催化集的交换遗传几乎肯定在目前在所有 6700 种原核生物中发现的小分子自催化集的早期进化和系统发育中发挥了重要作用 (11,12)。不可约自催化集可以通过计算来识别。因此,现在可以对这种新陈代谢的进化进行详细研究。
在分子多样性和复杂性逐渐增加的宇宙化学演化中,必然涌现出分子繁殖(复制)的一级相变。在宇宙形成的最早期,基本粒子、夸克、胶子、电子、正电子。随着宇宙冷却,强子形成。然后形成了第一种元素:氢、铍。后来,在超新星中,其剩余的 98 种稳定元素形成了(30)。然后是简单分子,之后是越来越复杂的分子出现,遵循从简单到低多样性向上越来越复杂的的模式。分子的多样性、原子复杂性以及它们之间的潜在反应可能性增加了(31)。50 亿年前与我们所在的太阳系一齐形成的默奇森陨石(The Murchison meteorite)显示,其拥有数十万种分子种类以及它们之间的潜在反应可能性(32)。
本文中讨论的理论预测,当分子种类 M 和它们之间的反应 R 足够多样化的情况下,当 R/M 比值足够大时,集体自催化集将作为一级相变出现。
关于涌现的集体自催化集(RAF)理论已经很成熟(15,16,25,27,28,29)。我们在这里将 RAF 理论与另一个独立理论 TAP (33,34,35) 结合起来,新的理论可以解释不断演化的宇宙中分子物种多样性日益增加的现象。
2.6 The TAP Equation TAP 方程
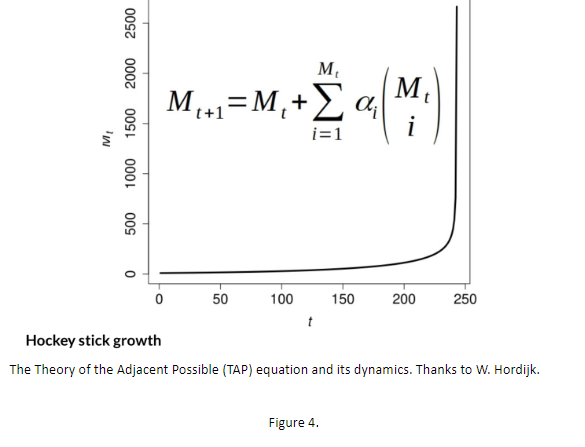
这个简单方程的动力学性质现在已经通过数值和定理得到了很好的研究(33,34,35)。这些特性是显著的。该系统是一个离散的动力系统,其中分子可以相互结合形成新的分子。如果当前分子数为 Mt,则对于在 Mt 中选择的每个大小为 i 的子集(Mt 选择 i),该方程将它们组合起来以概率 alphai 创建一个新分子,其中 0 < alpha < 1.0。
如果系统从少量起始分子开始,每个分子都可以与自身或其他分子的副本结合形成新分子,随着时间的推移,分子种类的数量缓慢增加,然后双曲线向上爆炸,并在有限时间内达到无穷大,(33,34 35),图 4。TAP 过程是宇宙化学多样性不断增加的粗略模型,(31,32)。
图4. 临近可能性理论方程及其动力学。|来源:W. Hordijk
我们现在将 TAP 和 RAF 联合起来。该联合理论希望将预期(必然)出现的集体自催化系统解释为不断演化的宇宙中的一级相变。简单的步骤是让 TAP 中的每个分子以固定概率 Pcat 随机催化 TAP 中的每个反应。以此理论解释技术进化的第一篇文章刚刚发表(36)。随着时间的推移,物质的多样性增加,从而出现一级相变到集体自催化集的概率上升到1.0(36)。
这个TAP-RAF 联合理论证明了这样一个基本事实。即,在宇宙的化学演化过程中,分子多样性通过一些类似于 TAP 过程的过程而增加。当这种情况发生时,分子的复杂性增加,因此反应的数量增加,反应的比率R与分子M、R/M的比率也增加。但这里,催化集中的分子M和反应集的分子R,实际上都来自于同一组候选分子集合。因此,M对于任何粗略的催化概率Pcat,按照幂律或其他方式均匀分配在分子之间,在某个时间点会出现一级相变,分子复制在不断演化的宇宙中预期会必然出现。
活力(Élan Vital) 的自然出现——进化中的生命
任何这样的非平衡可再生分子反应系统都是实现催化闭合的康德式整体。此外,充当催化剂的分子本身就是它们催化的特定反应中能量释放的边界约束条件。因此,系统构建了自己的关于能量释放的边界约束条件,该能量释放也构建了系统自身的边界约束条件,从而使系统实现了约束闭合。(在这基础上)我们添加某种形式的外壳,例如,在热液喷口的“小口袋”中,或者更好的的情况是在脂质体中,系统中的脂质膜也是由这同一化学催化系统中合成的(37,38)。
这种新构建的实现了上述四种闭合(即康德式整体、催化闭合、约束闭合和空间闭合)的组合整体,就是Bergson所谓的神秘的活力(Bergson’s mysterious élan vital)(39),这个词在这里完全被祛魅了。这四个条件构成了生命,生命是在不断演化的宇宙中随着可预期(必然发生)的相变而出现的。
我们注意到 R. Hazen 及其同事最近提出的关于非生命世界功能多样性出现的新法则理论的相似之处 (40)。作者考虑了一组稳定的实体,例如多种分子,以及它们之间的稳定变换,例如分子之间的化学反应,并指出复杂的系统,例如高度多样性的非生命矿物质的形成。“功能”是针对矿物质的特征而定义的。这些作者没有考虑康德式整体的出现,即实现催化闭合和约束闭合的集体自催化集,以及可进化生命的出现。在这里,“功能”的定义是部分如何演化以维持不断演化的整体。
3. 第二个奇迹:生物圈的演化是一种传播性的、不可推导式的构造,而不是必然的演绎。不存在定律。演化是不断创造的过程。
所有经典物理学,即牛顿物理学,都符合在牛顿范式:(1)陈述相关变量,例如位置和动量。(2)以微分形式陈述连接相关变量的运动定律。牛顿运动三定律就是一个例子。(3)定义边界条件。这些定义了相关变量值的所有可能组合的相空间。(4)说出初始条件。(5)对运动方程进行积分,以获得系统相空间中必然确定的单个轨迹 (41,42,43)。
量子力学中的牛顿范式没有改变。对薛定谔波动方程进行积分以获得概率分布的蕴含轨迹。然后进行通常在本体论意义上的随机测量(42,43)。
必须事先指定相空间,这是整个牛顿范式的基础,因此也是所有现代物理学的基础(42,43)。接下来我们表明,康德式整体不断发展的生物圈持续创造了无法提前推断或确定的新相空间。因此,整个牛顿范式(在面对生命问题时)崩溃了。不断演变的生物圈不能仅用物理学来解释,诉诸“功能”是必要的。
1.一旦我们定义了康德式整体,对部分“功能”的非循环定义就很清楚了。“部分的功能”是用于维持整体因果后果的子集。心脏的功能是泵血,而不是发出心音或使心包囊中的水摇动。
2. 选择是作用于康德式整体的整体水平上的,而不是作用于它的各个部分。选择并不直接选择泵血效率更高的心脏,是因为遗传了这种改良心脏的生物更有可能产生后代,因此改良的心脏是被间接选择的。
3. 因为“部分的功能”是维持整体因果属性的子集,所以同一部分的功能可以改变。同一部分的一些新的、未使用的因果属性子集可以用来维持整体。这些被称为达尔文预适应,或者被古尔德和维巴称为扩展适应(44)。例如,一些恐龙为了调节体温而进化出的鳞片,后来进化成了鸟类的飞羽。其他例子包括选择正常的酶变成透明的晶状体蛋白。一个极好的例子是从肺鱼的肺部进化出鱼鳔(43)。
4. 这种扩展的一个显著且基本的特征是它们无法被推理得出。考虑一个假设的例子。发动机缸体可以用作镇纸。同样的发动机缸体有尖角,可以用来敲开椰子。但从发动机缸体被用作镇纸的事实来看,并不能推断出“这个物体可以用来敲开椰子”,因为香蕉皮也可能被用作镇纸。
5. 如果同一物体的这种新用途是“应急拼凑的”,那么就不存在对于这种应急胡乱拼凑结构的演绎理论。
6. 真正深刻的含义是,这种不可推论的临时调整是功能新颖性和生物圈开放式进化的源泉。生物圈的进化是一种不可演绎的构造,而不是必然的演绎。没有必然的规律支配生物圈的演化,(42,43)。
7. 进一步的含义同样深刻。我们无法列出发动机缸体单独或与其他东西一起使用的所有用途。我们也无法列出螺丝刀单独或与其他东西一起使用的所有用途。因此,我们不能使用集合论或任何基于集合论的数学:集合论的第一公理是外延性公理,(45):“两个集合相等当且仅当它们包含相同的 11 个成员”。但我们无法证明发动机缸体的未列入清单的用途与螺丝刀的未列入清单的用途相同。更多选择公理也失败了。其含义是巨大的:我们不能使用任何基于集合论的数学——基本上全部数学——来推断生物圈的未来演化。
8. 这意味着(对于生命生成与演化)的集合没有并集和交集。对于生命生成与演化过程,我们无法构建一阶逻辑,无法使用 Russell 理论进行定性推演,(43, 46),无法使用皮亚诺公理系统为其定量,(43, 47),无法写出方程式。无法使用实数、实线、歧管的概念,无法使用拓扑工具,也无法写出流形上的方程,包括方程的积分。
9. 整个牛顿范式是所有物理学的基础,需要一个预先规定的相空间。但我们既不能推论也不能预测进化生物圈的进化相空间(42,43)。进化论超越了牛顿范式。
10. 我们无法仅用物理学来解释不断演变的生物圈。心脏凭借其泵血功能而进化。可遗传变异的自然选择作用于康德的整体,而不是直接作用于其部分。这种选择就是向下因果关系。这里,解释性箭头指向上方。
11. 许多人的梦想——强烈的还原论——失败了。如果刻在著名T恤上的最终理论是包括生物圈演化的推论,那么就没有最终理论。
12. 海森堡对不确定性关系的论证要求我们放弃经典物理学(48)。
13. 如果我们必须放弃生物进化的集合论,这意味着什么?我们几乎不知道。
14. 我们现在进入了科学的第三次范式转变(43)。新的基本问题无疑包括:随着生命之花的蓬勃绽放,不断演变的生物圈会如何应对呢?首先,它通过遗传变异和自然选择,或者遗传漂变的方式,来创造出新的可能性。这些可能性如同不断涌现的“泡沫”,代表着生命体能在某段时间内共存的新的方式。生物圈会抓住这些新方式,使它们得以固化并继续向前演进。
1. 我们正在发现越来越多的系外行星(49,50),并在这些行星的大气层中寻找生命的证据(50),本文讨论的结果展示了一个强有力的新希望。如果我们确定所有 6700 种原核生物中的小分子集体自催化集确实可以在体外进行化学繁殖(复制),那么我们就可以在系外行星的大气层中寻找此类小分子集体自催化集的证据。这些集合可能是宇宙中最基本的生命形式。给定大气中一组已识别的小分子,就可以通过计算来测试和判断该集合是否可以作为小分子集体自催化集发挥作用,这正是 Xavier 等所做的工作(11,12)。如果他们的结果被证实,我们可能会说在体外获得了生命。
2. 目前人工创建高多样性小分子文库是可行的。例如,Ott 进行的 Miller Urey 实验从仅四种分子开始,经过一个月的相互作用,产生了数千种小分子,并通过质谱法进行了鉴定 (51)。我们现在可以问,在这样的系统中是否可以出现小分子集体自催化集。这是一个生死未卜(“Go or No Go”)的实验。如果结果为“否”,则说明该理论可能是错误的。
3. 如果答案为“是”,我们就可以开始设想超越小分子集体自催化集的可测试途径,这些组份为肽RNA自催化集的代谢网络,它们可以共同进化为新的康德式整体。接下来,这些集体自催化网络可能会进一步演化出模板复制,甚至进化出基于遗传编码的复制(52,53,54)。当然,进一步还需要许多真实的实验来验证。
4. 我们由此可以开始设想从最早的小分子康德式整体到原核生物的出现的可测试途径。从这些发展到真核生物可能是一个漫长的步骤,但多细胞生物出现了六次或更多次。复杂生命的出现真的那么不可能吗?我们不知道。也许不是。
我们在地球上各民族的所有创世神话中寻找生命的源头,也许可以追溯到50万年前的尼安德特人。生命起源问题作为一个科学问题是随着帕斯特的主张而出现的:生命只来自生命(55)。
自 Haldane 和 Oparin (56,57) 以及 1953 年著名的 Miller Urey 实验 (58) 以来,实验工作一直在进行。基于生命必须以多核苷酸模板复制为基础的信念,已经进行了大量的努力(4,5,6)。然而迄今为止,人们尚未在这条合理途径上发现分子复制的案例。集体自催化集涌现的概念于 1971 年首次提出(14)。这种 DNA、RNA 和肽的复制化学组是使用进化的聚合物序列设计的。Xavier 现在在所有 6700 种原核生物中发现了不含 DNA、RNA 或肽聚合物的小分子集体自催化集 (11,12)。这强烈表明(虽然并不能证明),这样的系统可能是宇宙中分子繁殖(复制)的最早形式。
本文通过完善的数学理论表明,这种系统可以作为分子多样性和原子复杂性增加的宇宙中的一级相变而出现。如果这是正确的,正如我们所说,生命的出现是不断演化的宇宙中可预期(必然发生)的相变。自牛顿以来,基础科学一直依赖于强大的牛顿范式。该范式需要相空间中相关变量的所有值的预先设定且已知。但生命体是实现了催化闭合、约束闭合和空间闭合的康德式整体。令人惊讶的是,我们无法使用任何基于集合论的数学(即,似乎目前所有的数学)来推断生物圈不断创造性的进化。生命是宇宙中一个令人期待的奇迹,其形成的方式实际上是无限的。
20 世纪见证了原子时代、蘑菇云和相互毁灭的出现。对于哥德尔来说,20 世纪也见证了确定性的终结 (59)。在 21 世纪的第一个四分之一世纪,我们却几乎看不到我们所属的生物圈是否还具有惊人的蓬勃发展的创造力。我们确实属于自然,而不是凌驾于自然之上。
-
Schrödinger, E. (1992) What is life?: With mind and matter and autobiographical sketches. Cambridge University Press.
-
Hordijk, W. (2013) Autocatalytic sets: From the origin of life to the economy. BioScience 63, no. 11, 877-881.
-
Farmer, J. D., Kauffman, S. A. and Packard, N. H. (1986) Autocatalytic Replication of Polymers. Physica D 2, 50-67.
-
Orgel, L. E. The origin of life on the earth. (1994) Scientific American 271, no. 4 76-83
-
Paul, N. and G. F. Joyce. (2004) Minimal self-replicating systems. Current opinion in chemical biology 8, no. 6, 634-639.
-
Szostak, J. W. (2012) The eightfold path to non-enzymatic RNA replication. Journal of Systems Chemistry 3, 1-14.
-
Sievers, D. and von Kiedrowski, G. (1998) Self‐replication of hexadeoxynucleotide analogues: autocatalysis versus cross‐catalysis. Chemistry–A European Journal 4, no. 4, 629-641.
-
Vaidya, N., Manapat, M. L., Chen, I. A., Xulvi-Brunet, R., Hayden, E. J. and Lehman, N. E. (2012) Spontaneous network formation among cooperative RNA replicators. Nature 491, no. 7422, 72-77.
-
Wagner, N. and Ashkenasy, G. (2015) How catalytic order drives the complexification of molecular replication networks. Israel Journal of Chemistry 55, no. 8, 880-890.
-
Lancet, D., Zidovetzki, R. and Markovitch, O. (2018) Systems protobiology: origin of life in lipid catalytic networks. Journal of The Royal Society Interface 15, no. 144,
-
Xaiver, J., Hordijk, W., Kauffman, S. A., Steel, M., Martin, W. (2020), Autocatalytic chemical networks at the origin of metabolism Proc Roy Soc B, 287: 20192377. https:/doi.org/10.1098/rspb.2019.2377
-
Xavier, J. and Kauffman, S. A., (2022). Small-molecule autocatalytic networks are universal metabolic fossils. Proc Roy Soc A. 380:20210244
-
Kant, I. (2017) Critique of Judgement by Immanuel Kant-Delphi Classics (Illustrated). Vol. 11. Delphi Classics.
-
Kauffman, S. A. (1971). Cellular Homeostasis, Epigenesis, and Replication in Randomly Aggregated Macromolecular Systems. Journal of Cybernetics 1, 71- 96.
-
Kauffman, S. A. (1986). Autocatalytic Sets of Proteins. J Theor Biol 119, 1-24.
-
Hordijk, W., Hein, J. and Mike Steel. (2010) Autocatalytic sets and the origin of life. Entropy 12, no. 7 1733-1742.
-
Montévil, M. and Mossio, M. (2015) Biological organisation as closure of constraints. Journal of theoretical biology 372 179-191.
-
Atkins, P.W. (1984) The Second Law, Scientific American Library, NY.
-
Kauffman, S. A. (2000) Investigations, Oxford University Press, NY.
-
Von Neumann, J. (1966) Theory of self-reproducing automata. Arthur W. Burks, ed. University of Illinois Press.
-
Davies, P. (2019) The Demon in the machine: How hidden webs of information are solving the mystery of life. University of Chicago Press.
-
Margulis, L. (1971) Symbiosis and evolution. Scientific American225, no. 2, 48-61.
-
19Margulis , L. and Sagan, D. (2000) What is Life? University of California Press.
-
Campbell, N. A., Williamson, B., Heyden, R. (2006) Biology: Exploring Life. Boston, Massachusetts: Pearson Prentice Hall.
-
Hordijk, W., and Steel, M. (2018) Autocatalytic networks at the basis of life’s origin and organization. Life 8, no. 4, 62.
-
Erdős, P., and Rényi, A. (1960) On the evolution of random graphs. Publ. math. inst. hung. acad. sci 5, no. 1, 17-60.
-
Hordijk, W., and Steel, M. (2012) Autocatalytic sets extended: Dynamics, inhibition, and a generalization. Journal of Systems Chemistry 3, no. 1, 1-12.
-
Hordijk, W, Steel, M., Kauffman, S. A. (2012).The structure of autocatalytic sets: evolvability, enablement, and emergence. Acta Biotheoretica, Vol 60, Issue 4, 379-
-
Vassas. V., Fernando, C., Santos, M., Kauffman, S., Szathmary, E. (2012) Evolution Before Genes. Biology Direct. http://www.biology-direct.com/content/7/1/1
-
Puy, D. and Signore, M. (2002) From nuclei to atoms and molecules: the chemical history of the early universe. New Astronomy Reviews 46, no. 11, 709-723.
-
Kauffman, S. A., Jelenfi, D. Vattay, G. (2020) Theory of chemical evolution of molecule compositions in the universe, in the Miller-Urey experiment and the mass distribution of interstellar and intergalactic molecules J. Theor. Biol. Vol 486, https://doi.org/10.1016/j.jtbi.2019.110097.
-
Schmitt-Kopplin, P., Gabelica, Z., Gougeon, R., Hertkorn, N. (2010) High molecular diversity of extraterrestrial organic matter in Murchison meteorite revealed 40 years after its fall. Proceedings of the National Academy of Sciences 107, no. 7, 2763-276.
-
Steel,M., Hordijk W., Kauffman, S. A. (2020), Dynamics of a birth–death process based on combinatorial innovation, J. Theor. Biol 491, 11018.
-
Cortês, M., Kauffman, S. A., Liddle, A. R., Smolin L, Biocosmology and the theory ofthe adjacent possible. (2022) https://www.biocosmology.earth arXiv https://arxiv.org/abs/2204.14115
-
Koppl, R., Gatti, R., Deveraux, A., Fath, B., Herriot J., Hordijk, W., Kauffman S. A., Ulanowitcz, R., Valverde, S. (2023) Explaining Technology, Cambridge University Press, UK.
-
Hordijk, W. Koppl, R., Kauffman, S. A. (2023) Emergence of Autocatalytic Sets in a Simple Model of Technological Evolution, Journal of Evolutionary Economics, https://doi.org/10.1007/s00191-023-00838-2
-
Lane, N. (2015) Vital Question: Energy, Evolution, and the Origins of Complex Life. WW Norton & Company.
-
Damer, B., and Deamer, D. (2020) The hot spring hypothesis for an origin of life. Astrobiology 20, no. 4, 429-452.
-
Bergson, H. (1907). Creative Evolution. East India Publishing Company, Ottawa,Canada, 2022.
-
Wong, M., Cleland, C., Arend D., Bartlett, S., Cleaves, H., Demarest, H., Prabhu, A., Lunine, J. and Hazen R. (2023) On the roles of function and selection in evolving systems. Proceedings of the National Academy of Sciences 120, no. 43: e2310223120.
-
Smolin, L. (2013) Time reborn: From the crisis in physics to the future of the universe. HMH.2021
-
Kauffman, S. A. and Roli, A. (2021) The world is not a theorem. Entropy Vol 23, issue 11
-
Kauffman, S. A. and Roli, A. (2023) A third transition in science? Interface Focus13: https://doi.org/10.1098/rsfs.2022.0063
-
Gould, S. J., and Vrba, E. S. (1982) Exaptation—a missing term in the science of form. Paleobiology 8, no. 1, 4-15.
-
Fraenkel, A., Bar-Hillel, Y. and Levy, A. (1973) Foundations of set theory. Elsevier.
-
Grattan-Guinness, I. (2000) The search for mathematical roots, 1870-1940: logics, set theories and the foundations of mathematics from Cantor through Russell to Gödel. Princeton University Press.
-
Skolem, Th. (1955) Peano’s axioms and models of arithmetic. In Studies in Logic and the Foundations of Mathematics, vol. 16, pp. 1-14. Elsevier.
-
Cassidy, D. (1992) Heisenberg, uncertainty and the quantum revolution. Scientific American 266, no. 5, 106-113.
-
-
Krissansen-Totton, J., Thomson, M., Galloway, M., Fortney, J. (2022) Understanding planetary context to enable life detection on exoplanets and testing the Copernican Principle, Nature Astronomy, Vol 6, 189-198.
-
Wollrab, E., and Ott, A. (2018) A Miller–Urey broth mirrors the mass density distribution of all Beilstein indexed organic molecules. New Journal of Physics 20, no. 10 105003.
-
Lehman, N. E. and Kauffman, S. A, (2021), Constraint closure drove major transitions in the origins of life.Entropy, 23, 1, 105. https://doi.org/10.3390/e23010105
-
Carter C., Wills, P. (2017), Interdependence, Reflexivity, Fidelity, Impedence Matching, and the Evolution of Genetic Coding. Molecular Biology and Evolution, Vol 35, issue 2.
-
Kauffman, S. and Lehman, N. E. (2023) Mixed anhydrides at the intersection between peptide and RNA autocatalytic sets: evolution of biological coding, Journal of the Royal Society Interface. https://doi.org/10.1098/rsfs.2023.0009
-
Bordenave, G., Louis Pasteur (1822–1895). (2003) Microbes and infection 5, no. 6, 553-560.
-
Lazcano, A. (2016) Alexandr I. Oparin and the origin of life: a historical reassessment of the heterotrophic theory. Journal of molecular evolution 83, 214-222
-
Tirard, S. (2017) JBS Haldane and the origin of life. Journal of genetics 96, no. 5, 735-739.
-
Miller, S. and Urey, H. (1959) Organic Compound Synthesis on the Primitive Earth. Science Vol 130 (3370) 245–51.
-
Macintyre, A. (2011) The impact of Gödel’s incompleteness theorems on mathematics. Kurt Gödel and the foundations of mathematics: Horizons of Truth 3-25.
集智俱乐部联合西湖大学理学院及交叉科学中心讲席教授汤雷翰,国家蛋白质科学中心(北京)副研究员常乘、李杨,香港浸会大学助理教授唐乾元,北京大学前沿交叉学科研究院研究员林一瀚,中国科学院分子细胞科学卓越创新中心博士后唐诗婕,共同发起「生命复杂性:生命复杂系统的构成原理」读书会,从微观细胞尺度、介观组织器官尺度,到宏观人体尺度,梳理生命科学领域中的重要问题以及重要数据,希望促进统计物理、机器学习方法研究者和组学研究者之间的深度交流,建立跨学科合作关系,激发新的研究思路和合作项目。读书会从2024年8月6日开始,每周二晚19:00-21:00进行,持续时间预计10-12周。欢迎对这个生命科学、物理学、复杂科学、计算机科学深度交叉的前沿领域感兴趣的朋友加入!
6. 加入集智,一起复杂!