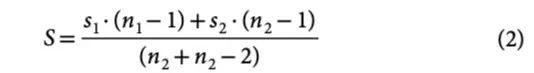人类文化与生物种群演化速度哪个更快?答案也许出乎意料:它们几乎是同速。2 月在 Nature Human Behaviour 发表的一篇论文揭示,文化和生物进化的过程和模式具有深度统一性。研究者通过使用时间序列分析法,得出大部分演化由某种稳定力或方向力在推动,并分别在一定程度上调节着不同性状的演化速率。通过文化选择,很多人工文化品的特征演变可以由“最优转移模型”来解释,而该模型又基于人类审美偏好中已知的种种心理偏差。
当今世界似乎充满变化,不仅新的文化和消费产品令人目接不暇,新的艺术实验、科技成果也层出不穷,并广泛渗入到所有人的生活日常。即使不追随时代潮流的人,也早已习惯了“旧的”迟早会被“新的”取代,就如每年总是一场场如期而至手机发布会上的一阵阵惊呼。
只有念旧的人们环顾自然或忆念往昔时,眼里还能依稀浮现出些许慢时光们的美好。
几乎所有人都已经达成了这样的共识:人类文明在加速发展,已经远远超出了自然演化的速度。一些研究者认为[2-5] ,这是因为思想在头脑中传播的速度,要比基因在身体中快得多。用道金斯的术语来说,这是一个模因(meme)主导基因、文化胜过自然的时代,在某些人类沙文主义者眼中,也是人类优越于其他物种的理由。顺应趋势,2016 年的世界地质大会上正式宣布,继更新世和全新世之后,地球已经进入一个全新的地质年代——“人类世”(Anthropocene)[1]。
这个全新纪元是以工业化和全球殖民的大加速为特征的,它意味着人类已经具备了干预星球进程的能力,并直接导向科幻爱好者和未来学家们所津津乐道的加速主义(Accelerationism)、超人类主义(Transhumanism)或奇点主义(Singularitarianism)。

然而,就如同人类曾普遍信奉万物有灵一样,以上浮光般的感受和论证有没有可能只是一种新的盲目信念?美国历史和社会学家刘易斯 · 芒福德(Lewis Mumford)曾在两卷本的技术理论专著《机器神话》中写道:
我们当今文明的先行者们,往往把他们特殊的机械技术成就和进步方式与那些毫无道理的道德优越感混为一谈……维多利亚时代就有种信仰,相信通过掌握机器,人类其他一切组织制度和习俗规约必然都能大大改进。当今的人类本该抵制、拒绝这种无根据的信仰。
诚如芒福德所言,如果没有足够的科学证据,即使感受再强烈,一种信念并不优于另一种信念,尤其在涉及如此宏大的议题时。
最近在 Nature 的 Human Behaviour 板块上发表的一篇论文,对自然和文化演化速率进行了定量研究。在《The pace of modern culture》这篇论文中,作者研究了流行音乐、汽车、小说等人类文化制品的进化动力学,并与有机生物种群演化进行了横向对比。
The pace of modern culture
https://www.nature.com/articles/s41562-019-0802-4
与普遍认为人类文化发展非常迅速的看法相反,研究表明现代文化的演化速度与许多动物种群相似。通过使用时间序列分析法,论文得出大部分现代文化是由某种稳定力或方向力,或二者共同作用而形成的,它们分别在一定程度上调节着不同文化特征的演化速率。通过文化选择,很多人工文化品的特征演变可以由“最优转移模型”来解释,而该模型又基于人类审美偏好中已知的种种心理偏差。
研究者们选取了流行音乐、小说、临床医学文献、汽车四种人工文化制品,与有机生物种群中的炭疽菌、红肩甲虫、丛林蜗牛、三刺鱼、中型雀等进行了对比。
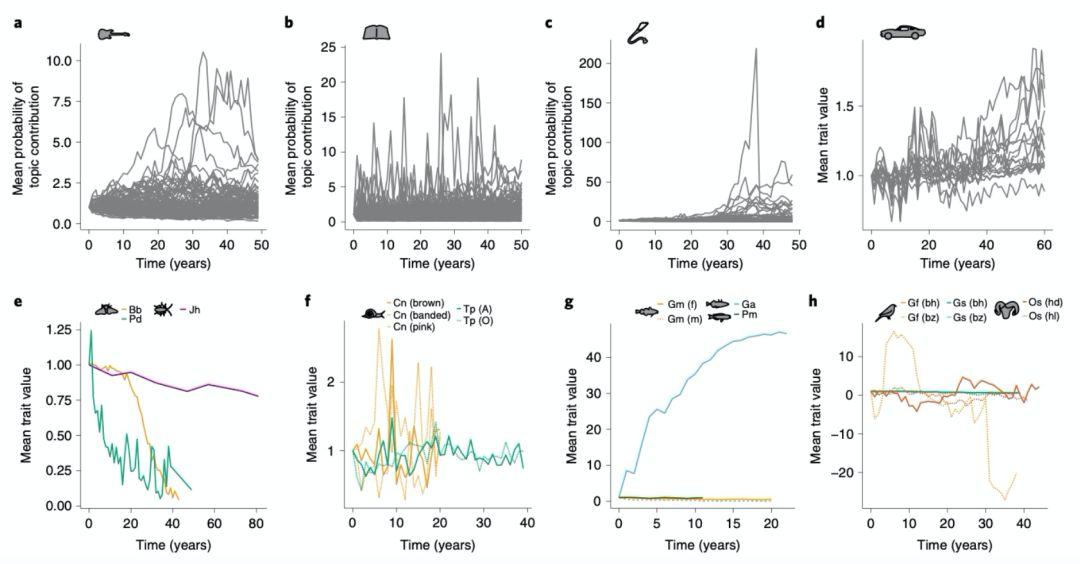
在图 1 中,可以看到对这些文化制品和种群所测定各种性状的原始演化轨迹。
但由于时间序列的分辨率不同,它们的演化速度不能简单通过观察来对比。为此研究者们选取了生物学中几种演化指标 [6,7]中的一种:Haldanes 速率,进行比较。这种指标将物种变异的性状当做原始资料,能够体现出种群内部变异情况。
在一个不断演化的种群中,对于具有某种性状的表现型 z1 和 z2,Haldane 速率(h)是时间间隔 I = t2 – t1 下、以标准差校正后的变化率,它代表了种群性状演化速度变化情况:
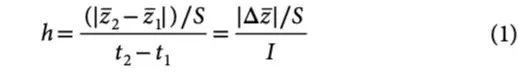
其中 S 是合并样本后标准差,由两个时间点的样本标准差 s1 和 s2 以及样本大小 n1 和 n2 计算得出:
在(1)中,ν = ∣Δz∣∕ S 是 h 的分子,代表性状 z 经样本标准差校正后的变化情况,即 Haldane 速率 h 则是 ν 在时间上的变化率。
图 2:流行音乐主题演化轨迹和对应的 Haldane 速率
可以看到,在五十年内,不仅音乐主题多样性在到一定高度后开始下降,从整体演化看,速度 h 除了在10-25区间短暂加速外也一直在降低。
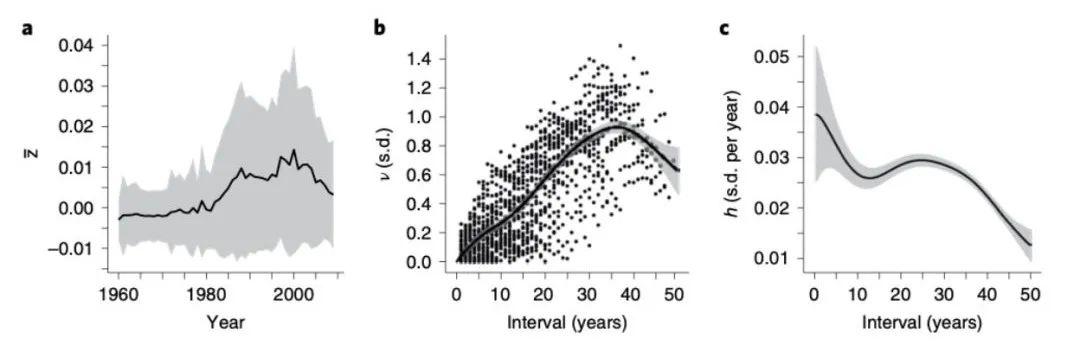
在使用以年为单位时间对输出数据进行归一化处理后,研究者们计算出了所有文化制品和有机生物种群的绝对 Haldanes 速率(即以年为单位的 h1),如图 3 所示:

图 3:文化和生物种群演化的绝对 Haldanes 速率
此前已有一些研究表明 [7-10,37],有机生物种群的进化速度会随着时间间隔增加而变缓,这与本文研究结论相符(图3e–h)。对比人工文化制品,从图 3a-d 曲线可以看到,它们和有机演化呈现出了相同的演化趋势。
演化速度随着时间间隔的增加而变慢,这意味着从长远来看,大多数性状特征比每年直接观察看到的更为保守。例如,在流行音乐的各种特征中,主题表现型的变化可能都在同一方向上。因此即使再经过五十年发展,平均偏移也只有 0.005个标准差。这可以解释为什么对所有艺术家的歌曲风格而言, 在排行榜上新的流行歌曲很少听起来是完全陌生的。
同样明显的是,不同有机生物种群的演化速度各有快慢(图3e–h)。若采取整数年为单位计算,某些动物的演化速率变动可能会超过一个数量级。
其中进化最快的动物通常会涉及遗传多态性(Polymorphisms),即存在两个或多个等位基因的表型。例如一种由自然选择定向驱动演化 [15] 的炭疽菌(Bb),每年以 h1 = 1.56 的速率演化。而外壳颜色多态性较大丛林蜗牛则以 h1 > 3 的演化。其中最快是一种三刺鱼(Ga),鳍甲以 h22 = 6.0 的速率快速变化(22 年平均)。
至于受到环境影响较大的连续性状,如其中一种达尔文雀(Gf&Gs),与多态性种群相比,演化速率 h1 则变化较低。
物种多态性种群处在几个等位基因的强大遗传控制之下,这意味着它们的 Haldanes 速率是真实演化速率大小。相比之下,连续性状的变化几乎大部都是由表型可塑性(phenotypic plasticity)引起的 [20,21] [16,17,18,22,23](其中一种方式是表观遗传),因此表型变化快慢可以视作演化速率的上限。
这样在纳入多态性考量后,为了对比人类文化和有机种群的演化速度,研究者们分别计算了四种人类文化制品的 h1 分布上限:发现它们均远小于有机性状的分布上限。并发现即使排除多态性的影响,文化和有机性状分布也处于相同的数量级——在每年间隔下 0.1 < h1 <1.0 s.d。
也就是说,文化和生物的演化速度令人惊讶地相似,并且有些生物性状的演化速度比观察到的任何文化特征都要快得多。
虽然文化和生物进化速度处于同一数量级,但如果考虑不同时间间隔,会发现绝对 Haldane 速率(h1)和25年平均速率(h25)还是有很大不同。前者反映了性状的短期变化,后者则代表族群的长期变化。
例如以隔年计算的 h1(图3a–d)而言,汽车发展速度比小说快 1.3 倍,比流行音乐和临床医学文献分别快3.6和8.1倍。但如果考虑25年间隔的h25,顺位虽然保持不变,数值差异却变得小了。在图 4a-b 中展示了文化和生物性状(一种蓝鸟),取对数后的 h1 和 h25 分布对比情况。
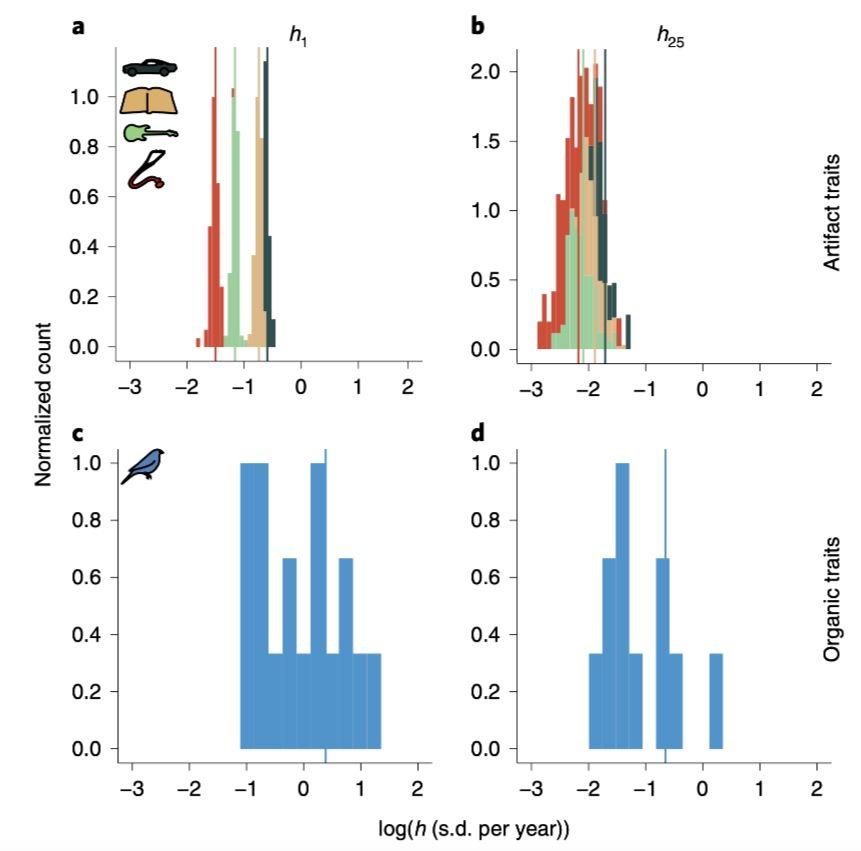
图 4:取对数后两种 Haldane 速率分布对比,垂线是中位数
可以看到,在长期而言不同性状变化会有很大差异:有些变化更快了,而另一些则更为保守。那么如何解释性状这种长期变化产生的差异呢?
考察一个有性生殖种群,新变异体产生的比率是有效种群大小、突变率以及选择形式和数量的函数 [24] [13,25-27]。
研究者们首先观察到,在没有任何其他力量的情况下,性状变化将呈现无偏随机游走(URW):下一代特征的预期值等于当前值。
而如果受到某种稳定力(stabilizing forces)的作用,性状将表现出的回归均值特征,具有持久性。若受到的是方向力(directional forces)影响,则会产生一个有偏向性的趋势 [28-30]。
这样的作用力模式为长期演化速度变化提供了两种解释:
-
保守性状受到稳定力的影响,快速演化性状则受影响较小。这意味着性状会因后者随机游走而进化。
-
保守性状也可能随机游走(但每个时长变化很小),而快速演化性状则由方向力驱动。
在以上情况并假设可加性下,无偏随机游走、均值回归和方向力驱动,就共同作用性状导致了物种长期演化速度的变化。据此可以将对演化的作用模式分为四种:
-
无偏随机游走(Unbiased random walk,URW),保守、开放性状均随机
-
有偏随机游走(Biased random walk,BRW),保守-随机,开放-方向力
-
无偏均值回归(Unbiased mean-reverting,UMR),保守-稳定力
-
有偏均值回归(Biased mean-reverting,BMR),保守-稳定力,开放-方向力
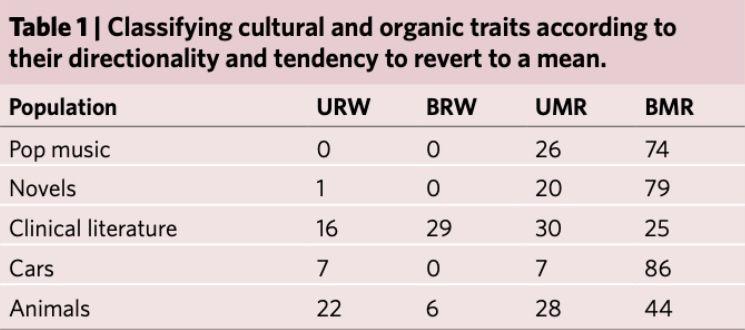
在 URW 和均值回归过程,演化速率都会随着时间间隔下降。研究者发现只有 0% 至 16% 的文化特质和 22% 的有机性状是无偏随机游走(URW)。而大约 25%-86% 的文化特征和 44% 的有机性状均表现出方向性和 BMR 动力学特性。这说明文化和有机种群都处在一定方向力作用下,同时由某种稳定力不断将自身特征值推向平均而演化。
表 1:不同人工文化品和动物种群中四项过程的百分占比。其中动物的 URW 最大,表示包含基因突变的生物演化更随机:
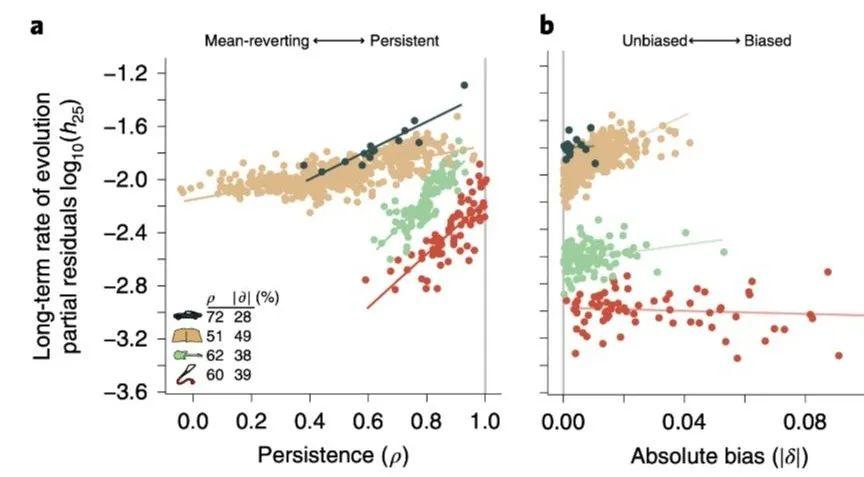
为了区分均值回归和一般随机游走的作用,研究者还分离变量进行了更细致的数学建模分析,将特定种群的长期演化率(h25)用方向力的偏向性(δ)和稳定力的持久性(ρ)测量。经过计算,二者可以解释大约 85%的统计显著差异。并且持久性于种群演化作用的影响平均三倍于偏向性。尤其对于人工文化制品,某些特征非常保守是因为它们受到特别强大的稳定力的约束,虽然定向力选择同时也在起作用。结果如图5所示。有趣的是,偏向性最高的是小说,而非作为科技产品的汽车,并且小说几乎是在保守和偏向之间取得了平衡。
图5:人工文化品在 h25下的持久性和偏向性(斜率),彩线为最小二乘拟合
那么,推动这些人工文化品演化背后的力量究竟是什么呢?
研究者认为,对应自然选择,其背后成因就是文化选择:由生产者(制造文化产品的人)、裁定者(分配和出售的人)、消费者(购买和使用的人)组成。与随机突变下自然选择不同的是,通过文化选择,某一种文化特征可以比另一种更容易获得和传播,其原因包括各种可能的传播偏好[3,13,14] 和文化吸引子[31-36]。
后者包括人类文化所依仗的一些心理能力,例如心智理论(Theory of mind)、生物归类(Folk biology)、数字能力、人脸识别、朴素力学(Naive mechanics)、族群分类(Folk sociology)等[32]。
对于传播偏好则有两种需要区分。一种是倾向于传播容易记住的模因。另一个是倾向于传播审美愉悦的模因。
在 2007 年曾有研究者通过从大量有机种群中收集的差异进行点估计,对宏观进化模型进行了拟合[12],认为演化数据可以用“最优转移模型”(shifting-optimum model)得到最好的解释。在该模型中,一个分散跨越了几代到1000万代时间尺度的表型,最核心的特征是在适应度最优的某个固定范围变动下演化,这与本研究结论一致。
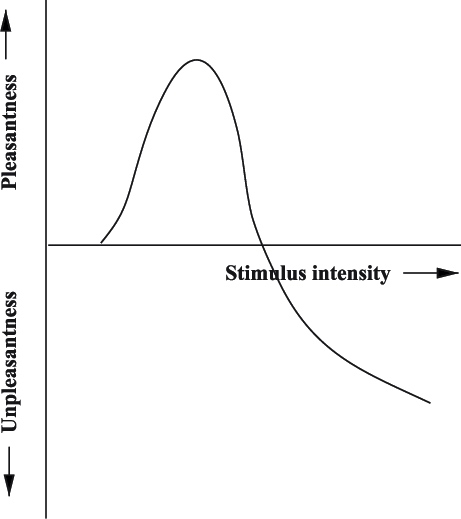
对文化选择而言,其中可能产生最佳转变的一种心理机制是,选择方在任何时候是否都持续偏向中等程度的新颖性,如享乐满意度的 Wundt-Berlyne 曲线[10]、心流曲线,以及我们之前介绍过的最优学习率和最佳审美配比。
我们可以看到,恰恰是每个人的审美和判断聚集起来,最终导致了整体文化的演变。
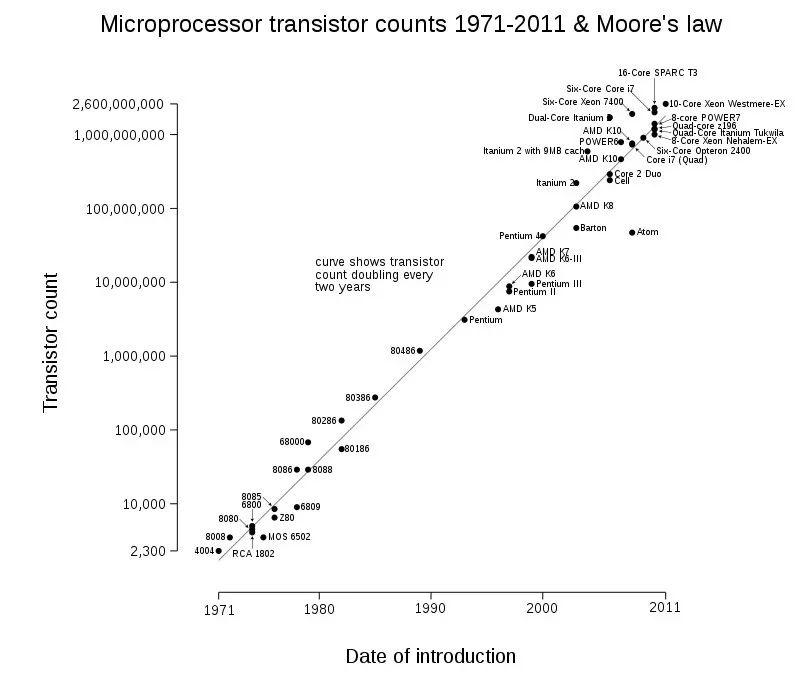
在本文中有一个很关键的遗留问题,即没有对更多具备技术特性的文化产品进行研究,尤其是基于互联网和计算产业的科技产品。
例如在计算机产业中存在的著名的摩尔定律。由于它不是基于性状分布的演化速度,因此不适用本文方法直接进行研究对比。但即使如此,最大晶体管密度24个月翻倍的速度,依然意味着计算机种群发展速度可能比最快的动物种群发展速度还快。
换句话说,在本文研究中,如果我们拥有的是现代汽车的计算能力而不是马力的数据,那么我们可能会发现汽车性能的一个维度甚至超过了三刺鱼的鳍甲。因此单单计算能力的提升,是否就意味着科技本身在加速演变中,这依然是一个值得深思的问题。这不仅需要在视角和概念上进行厘清,还需要更加准确和多维指标对科技发展进行衡量、以及分详细析科技对社会影响的关系。
例如,复杂系统科学中心圣塔菲研究所前所长杰弗里• 韦斯特(Geoffrey West),就认为在地球文明中加速发展的是城市,而非人类或人类文化。后者只不过是这种新型生物体中的细胞和能量。在他看来,导致城市超指数发展的是一切社会创新和财富创造活动,并让城市在越来越短的周期内面临“创新-崩溃循环”。地球真正所处时代是城市纪,而非人类纪。
因此,从本项研究我们还很难判断,当代技术是否导致了人类社会加速发展,是否让人类已经超越了自然生物的演化速度,亦或如生物演化一样只是一种局部时间的爆发,在长期演变看,实际与自然演化依然处在同一个数量级。
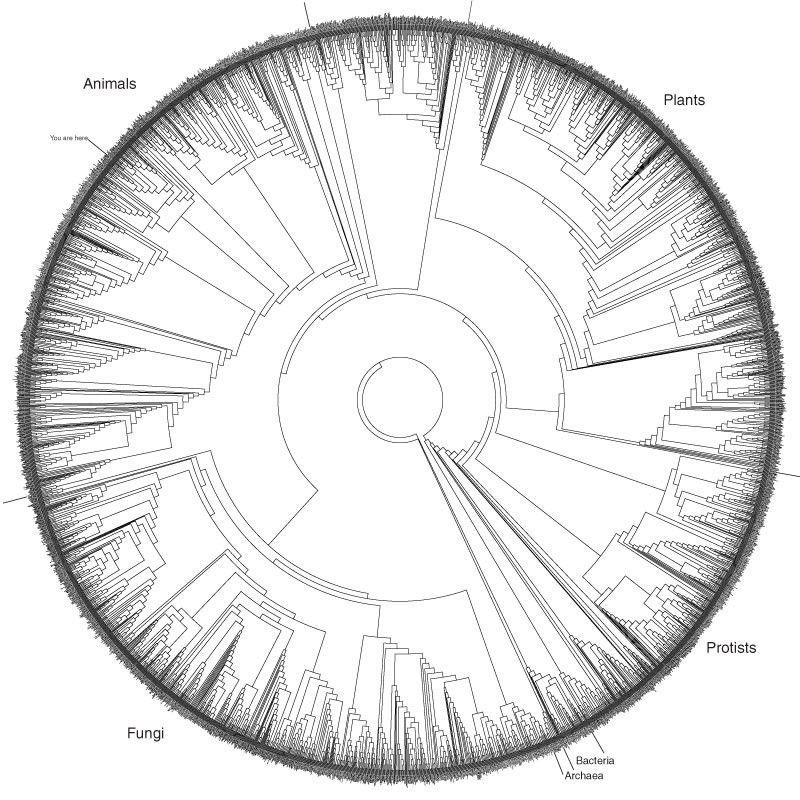
图8:系统生物学家 David Hillis 设计的演化圆盘,每个物种都在平等演化
也许问题的答案还在于你所处的视角,若以自然眼光看,人类依然是亿万种生物中的一种,也不是唯一的智能,没有脱离自然,更不可能离开宇宙。要想真正搞清楚问题,就不能盲目自大。就如技术本质也源于自然一样,对这个问题的回答,一种观点或信念没有意义,我们必须回到客观自然规律探索本身。
图9:装置艺术「最后的傀儡实验」,在处于顶点的赛博文明毁灭后,大自然重新开始生生不息
在“盖亚假说”中,地球就是一个巨大的生命有机体,在时刻调节着自己,就如希腊神话中那位产生诸神和一切生灵的大地女神。她让生命一时繁荣昌盛,也可能洗刷衰败的地表文明。
不管答案怎样,未来身处何方,能冷静看着人类并负责的,最终唯有人类自己。
[1] https://www.theguardian.com/environment/2016/aug/29/
declare-anthropocene-epoch-experts-urge-geological-congress-human-impact-earth
[2] Cronk, L. Intelligent design in cultural evolution. Behav. Brain Sci. 29, 352–353 (2006).
[3] Richerson, P. J., Boyd, R. & Henrich, J. Gene–culture coevolution in the age of genomics. Proc. Natl Acad. Sci. USA 107, 8985–8992 (2010).
[4] Mesoudi, A. Cultural Evolution: How Darwinian Theory Can Explain Human Culture and Synthesize the Social Sciences (Univ. Chicago Press, 2011).
[5] Perreault, C. The pace of cultural evolution. PLoS One 7, e45150 (2012)
[6] Haldane, J. B. S. Suggestions as to quantitative measurement of rates of evolution. Evolution 3, 51–56 (1949).
[7] Gingerich, P. D. Rates of evolution—effects of time and temporal scaling.
Science 222, 159–161 (1983).****
[8] Gingerich, P. D. Quantification and comparison of evolutionary rates.
Am. J. Sci. 293A, 453–478 (1993).
[9] Hendry, A. P. & Kinnison, M. T. The pace of modern life: measuring rates of contemporary microevolution. Evolution 53, 1637–1653 (1999).
[10] Kinnison, M. T. & Hendry, A. P. The pace of modern life II: from rates of contemporary microevolution to pattern and process. Genetica 112, 145–164 (2001).
[11] Rosen, C. & Zerner, H. Romanticism and Realism (Faber and Faber, 1984)
[12] Estes, S. & Arnold, S. J. Resolving the paradox of stasis: models with stabilizing selection explain evolutionary divergence on all timescales. Am.
Nat. 169, 227–244 (2007).
[13] Cavalli-Sforza, L. L. & Feldman, M. W . Cultural Transmission and Evolution: a Quantitative Approach. (Princeton Univ. Press, 1981).
[14] Boyd, R. & Richerson, P. J. Culture and the Evolutionary Process (Univ.
[15] Cook, L. M. The rise and fall of the carbonaria form of the peppered moth. Q. Rev. Biol. 78, 399–417 (2003).
[16] Grant, P. R. & Grant, B. R. Unpredictable evolution in a 30-year study of Darwin’s finches. Science 296, 707–711 (2002).
[17] Grant, P. R. & Grant, B. R. 40 Years of Evolution: Darwin’s Finches on Daphne Major Island (Princeton Univ. Press, 2014).
[18] Coltman, D. W . et al. Undesirable evolutionary consequences of trophy hunting. Nature 426, 655–658 (2003).
[19] Pigeon, G., Festa-Bianchet, M., Coltman, D. W . & Pelletier, F. Intense selective hunting leads to artificial evolution in horn size. Evol. Appl. 9, 521–530 (2016).
[20] Seeley, R. Intense natural selection caused a rapid morphological transition in a living marine snail. Proc. Natl Acad. Sci. USA 83, 6897–6901 (1986).
[21] Trussell, G. & Smith, L. Induced defenses in response to an invading crab predator: an explanation of historical and geographic phenotypic change.
Proc. Natl Acad. Sci. USA 97, 2123–2127 (2000).
[22] Olsen, E. et al. Maturation trends indicative of rapid evolution preceded the collapse of northern cod. Nature 428, 932–935 (2004).
[23] Carroll, S. et al. And the beak shall inherit—evolution in response to invasion. Ecol. Lett. 8, 944–951 (2005).
[24] Kimura, M. & Ohta, T. The average number of generations until fixation of a mutant gene in a finite population. Genetics 61, 763–771 (1969).
[25] Henrich, J. Demography and cultural evolution: how adaptive cultural processes can produce maladaptive losses: the Tasmanian case. Am. Antiq.
[26] Aoki, K., Lehmann, L. & Feldman, M. W . Rates of cultural change and patterns of cultural accumulation in stochastic models of social transmission. Theor. Popul. Biol. 79, 192–202 (2011).
[27] Nakahashi, W . The effect of cultural interaction on cumulative cultural evolution. J. Theor. Biol. 352, 6–15 (2014).
[28] Hunt, G. Fitting and comparing models of phyletic evolution: random walks and beyond. Paleobiology 32, 578–601 (2006).
[29] Hunt, G. The relative importance of directional change, random walks, and stasis in the evolution of fossil lineages. Proc. Natl Acad. Sci. USA 104, 18404–18408 (2007).
[30]Hunt, G. & Rabosky, D. L. Phenotypic evolution in fossil species: pattern and process. Annu. Rev. Earth Planet. Sci. 42, 421–441 (2014)
[31] Sperber, D. Explaining Culture (Blackwell, 1996).
[32] Sperber, D. & Hirschfeld, L. A. The cognitive foundations of cultural stability and diversity. Trends Cogn. Sci. 8, 40–46 (2004).
[33] Claidière, N. & Sperber, D. Imitation explains the propagation, not the stability of animal culture. Proc. R. Soc. B 277, 651–659 (2010).
[34] Claidière, N., Kirby, S. & Sperber, D. Effect of psychological bias separates cultural from biological evolution. Proc. Natl Acad. Sci. USA 109, E3526–E3526 (2012).
[35] Leroi, A. M., MacCallum, R. M., Mauch, M. & Burt, A. Reply to Claidière et al.: Role of psychological bias in evolution depends on the kind of culture. Proc. Natl Acad. Sci. USA 109, E3527–E3527 (2012).
[36] Cladière, N., Scott-Phillips, T. & Sperber, D. How Darwinian is cultural evolution? Philos. Trans. R. Soc. B 369, 1471–297 (2014).
[37] Gingerich, P. D. Rates of evolution. Annu. Rev. Ecol. Evol. Syst. 40, 657–675 (2009).
人类不平等的起源:为什么每一次技术变革都加剧了贫富分化
深度学习首次验证进化数学模型:趋同演化增加生物多样性
有关祖母假说、进化图模型以及生物进化的真相
进化新视角:基因之间的重复博弈如何影响生物进化?
加入集智,一起复杂!

集智俱乐部QQ群|877391004
商务合作及投稿转载|swarma@swarma.org
搜索公众号:集智俱乐部
加入“没有围墙的研究所”
让苹果砸得更猛烈些吧!
👇点击“阅读原文”,了解更多论文信息